Моторный блок
Содержание:
- Ссылки [ править ]
- Рефлекс на растяжение и мотонейронный пул
- Рефлекс на растяжение
- Способы объяснения «принципа величины»
- Единицы первого типа
- Вербовка (позвоночные) [ править ]
- Сокращение моторных единиц
- Внешние ссылки [ править ]
- Типы моторных единиц
- Типы двигательных единиц (позвоночные) [ править ]
- «Принцип величины» Хеннемана
- Мотонейронный пул
- Нарушения «принципа величины»
- Рефлекс «отряхивания лапы»
Ссылки [ править ]
- ^ a b Buchtal, F; Х. Шмальбрух (1 января 1980 г.). «Двигательная единица мышцы млекопитающих». Физиологические обзоры . 60 (1): 90–142. DOI10.1152 / Physrev.1980.60.1.90 . PMID 6766557 .
- Перейти ↑ Kandel, Eric (2013). Принципы неврологии, 5-е изд . Макгроу-Хилл, Нью-Йорк. п. 768. ISBN 978-0-07-139011-8.
- ^ Милнер-Brown HS, Stein RB, Yemm R (сентябрь 1973). «Упорядоченное задействование двигательных единиц человека во время произвольных изометрических сокращений» . J. Physiol . 230 (2): 359–70. DOI10.1113 / jphysiol.1973.sp010192 . PMC 1350367 . PMID 4350770 .
- Перейти ↑ Robinson R (февраль 2009 г.). «В мышцах млекопитающих проводка аксонов идет неожиданными путями» . PLOS Biol . 7 (2): e1000050. DOI10.1371 / journal.pbio.1000050 . PMC 2637923 . PMID 20076726 .
- ^ Фарина, Дарио; Merletti R; Енока Р.М. (2004). «Извлечение нейронных стратегий из поверхностной ЭМГ». Журнал прикладной физиологии . 96 (4): 1486–1495. DOI10.1152 / japplphysiol.01070.2003 . PMID 15016793 .
- ^ Spiegel KM .; Страттон Дж .; Берк-младший; Glendinning DS; Enoka RM (ноябрь 2012 г.). «Влияние возраста на оценку активации двигательных единиц в мышце руки человека». Экспериментальная физиология . 81 (5): 805–819. DOI10.1113 / expphysiol.1996.sp003978 . PMID 8889479 .
- ^ Де Лука, Карло; Уильям Дж. Форрест (декабрь 1972 г.). «Некоторые свойства потенциальных поездов действия двигательных единиц, записанные во время изометрических сокращений постоянной силы у человека». Кибернетик . 12 (3): 160–168. DOI10.1007 / bf00289169 . PMID 4712973 .
- ^ a b Burke RE, Levine DN, Tsairis P, Zajac FE (ноябрь 1973 г.). «Физиологические типы и гистохимические профили в двигательных единицах икроножной мышцы кошки» . J. Physiol . 234 (3): 723–48. DOI10.1113 / jphysiol.1973.sp010369 . PMC 1350696 . PMID 4148752 .
- ^ Collatos TC, Эджертон VR, Смит JL, Botterman BR (ноябрь 1977). «Сократительные свойства и состав волокон сгибателей и разгибателей локтевого сустава у кошек: значение для моторного контроля». J. Neurophysiol . 40 (6): 1292–300. DOI10,1152 / jn.1977.40.6.1292 . PMID 925731 .
- ^ Альтшулер Д .; Welch K .; Чо Б .; Welch D .; Lin A .; Дикинсон У .; Дикинсон М. (апрель 2010 г.). «Нервно-мышечный контроль кинематики взмахов крыльев у колибри Annas» . Журнал экспериментальной биологии . 213 (Pt 14): 2507–2514. DOI10,1242 / jeb.043497 . PMC 2892424 . PMID 20581280 .
- ^ a b Schiaffino S, Reggiani C (август 1994 г.). «Изоформы миозина в скелетных мышцах млекопитающих». J. Appl. Physiol . 77 (2): 493–501. DOI10.1152 / jappl.1994.77.2.493 . PMID 8002492 .
- ^ Б Caiozzo VJ, Baker МДж, Хуан К, Чжоу Н, У YZ, Baldwin КМ (сентябрь 2003 г.). «Полиморфизм тяжелой цепи одиночного волокна миозина: сколько паттернов и какие пропорции?». Являюсь. J. Physiol. Regul. Интегр. Комп. Physiol . 285 (3): R570–80. DOI10,1152 / ajpregu.00646.2002 . PMID 12738613 .
- Перейти ↑ Baldwin KM, Haddad F (январь 2001 г.). «Влияние различных парадигм активности и неактивности на экспрессию гена тяжелой цепи миозина в поперечно-полосатых мышцах». J. Appl. Physiol . 90 (1): 345–57. DOI10.1152 / jappl.2001.90.1.345 . PMID 11133928 .
- ^ Карпати, Джордж (2010). Расстройства произвольной мышцы . Издательство Кембриджского университета. п. 7. ISBN 9780521876292.упоминается Файнштейн, B; Линдегард, В; Найман, Э; Вольфарт, G (1955). «Морфологические исследования двигательных единиц в нормальных мышцах человека». Acta Anat (Базель) . 23 (2): 127–142. DOI10.1159 / 000140989 . PMID 14349537 .
Рефлекс на растяжение и мотонейронный пул
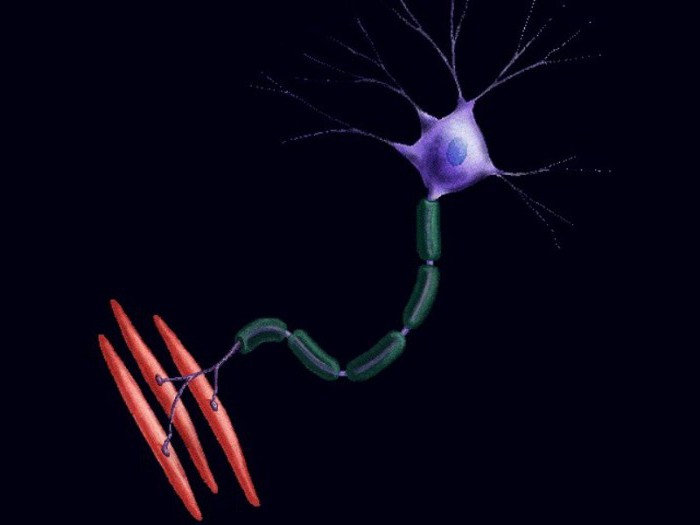
При растягивании любой мышцы первыми реагируют медленные волокна. Их нейроны генерируют разряды до десяти импульсов в секунду. Если мышцу продолжать растягивать, то частота генерируемых импульсов возрастет до пятидесяти. Это приведет к сокращению двигательных единиц третьего типа и увеличит силу мышцы в десять раз. При дальнейшем растяжении подключатся моторные волокна второго типа. Это преумножит силу мышцы еще в четыре-пять раз.
Двигательная мышечная единица управляется мотонейроном. Совокупность нервных клеток, входящих в состав одной мышцы, называется мотонейронный пул. В одном пуле могут одновременно находиться нейроны из разных, по качественным и количественным проявлениям, двигательных единиц. Из-за этого участки мышечных волокон включаются в работу не одновременно, а по мере того, как увеличивается напряжение и скорость нервных импульсов.
Рефлекс на растяжение
При растяжение мышцы первыми возбуждаются S-MH, но частота их разряда невелика (5—10 имп./с). По мере растяжения мышцы частота подачи импульсов S-MH растет и достигает 40—50 имп./с. При таком растяжении в работу включаются FR-MH. При подключении одной FR-MЕ сила возрастает примерно в 10 раз. Если растяжение продолжается, начинают импульсировать FF-MH , а значит подключаются FF-ME, каждая из которых дает прирост силы еще в 4—5 раз. Зависимость силы, которую развивает мышца от степени её растяжения или от длины называют мышечной характеристикой, которую можно отобразить на графике в виде кривой.
Способы объяснения «принципа величины»
С физической точки зрения, принцип Хеннемана можно объяснить тем, что у разных МН разное входное сопротивление. У небольших МН меньше площадь мембраны, а значит выше входное сопротивление.
Интересен способ измерения входного сопротивления. Его измеряют так: вводят микроэлектрод в клетку, пропускают через него ток и смотрят, как сильно изменился мембранный потенциал. Одинаковые синапсы в разных МН создают одинаковые синаптические токи, поскольку сопротивление самого синапса гораздо больше, чем входное сопротивление МН. У маленьких МН эти токи будут вызывать больший сдвиг мембранного потенциала и более сильную мембранную деполяризацию. Таким образом, сначала в работу включаются более мелкие МН.
Другая гипотеза, которая активно обсуждалась исследователями, объясняла «принцип величины», в связи с афферентами аI. Предполагалось, что афференты аI дают больше синаптических окончаний на маленьких МН и эти окончания лежат ближе к соме, а значит и эффективность синапсов выше. В процессе исследования этой идеи было получено множество количественных данных о связи мышечных афферентов и МН . Выяснилось, что одиночный афферент аI дает в пуле МН данной мышцы около 10 веточек, а каждая коллатераль образует до 200 синапсов, то есть всего афферент аI дает до 2000 синапсов. Например, в МН пуле трицепса 500—700 МН. На один МН в среднем приходится 2-4 синапса от одного афферентного волокна а I. С одним МН контактирует только одна коллатераль. Менделл и Хеннеман показали, что один афферент дает свои окончания достаточно диффузно по всему пулу, оканчиваясь на 90 % всех его МН и на 50 % мышцы синергиста. Таким образом можно предполагать, что вход от мышечных афферентов распределен по МН достаточно равномерно, так что именно собственные свойства МН определяют порядок их вовлечения.
Единицы первого типа
Двигательная единица первого типа или медленная неутомляемая, встречается чаще всего в крупных мышцах. Такие мотонейроны имеют низкий порог возбуждения и скорость проведения нервного импульса. Центральный отросток нервной клетки в своем терминальном отделе разветвляется и иннервирует небольшую группу волокон. Частота разрядов, поступающих к медленным двигательным единицам – от шести до десяти импульсов в секунду. Мотонейрон может поддерживать такой ритм в течение нескольких десятков минут.
Сила и скорость сокращения двигательных единиц первого типа в полтора раза меньше, чем у других типов моторных единиц. Причина этого – низкая скорость образования АТФ и медленных выходов ионов кальция на наружную мембрану клетки для связывания с тропонином.
Вербовка (позвоночные) [ править ]
Центральная нервная система отвечает за упорядоченное задействование мотонейронов , начиная с мельчайших моторных единиц. Принцип размера Хеннемана указывает на то, что двигательные единицы набираются от самых маленьких до самых больших в зависимости от размера нагрузки. При меньших нагрузках, требующих меньшего усилия, активируются медленные, малосиловые, устойчивые к усталости мышечные волокна, прежде чем задействуются быстрые, мощные и менее устойчивые к усталости мышечные волокна. Более крупные двигательные единицы обычно состоят из более быстрых мышечных волокон, которые генерируют более высокие силы.
Центральная нервная система имеет два различных способа управления силой, производимой мышцей посредством набора двигательных единиц: пространственное рекрутирование и временное рекрутирование. Пространственное вовлечение — это активация большего количества двигательных единиц для создания большей силы. Более крупные двигательные единицы сокращаются вместе с небольшими двигательными единицами до тех пор, пока не активируются все мышечные волокна в одной мышце, создавая, таким образом, максимальную мышечную силу. Рекрутирование временных моторных единиц или кодирование скорости связано с частотой активации сокращений мышечных волокон. Последовательная стимуляция волокон двигательных единиц от альфа-мотонейроназаставляет мышцу подергиваться чаще, пока подергивания не «срастутся» во времени. Это создает большую силу, чем единичные сокращения, за счет уменьшения интервала между стимуляциями для создания большей силы с тем же числом двигательных единиц.
С помощью электромиографии (ЭМГ) можно измерить нейронные стратегии мышечной активации. Порог разгонной силы относится к показателю размера двигательного нейрона, чтобы проверить принцип размера. Это проверяется путем определения порога включения двигательной единицы во время изометрического сокращения, при котором сила постепенно увеличивается. Двигательные единицы, задействованные при низкой силе (низкопороговые единицы), как правило, являются небольшими двигательными единицами, тогда как высокопороговые единицы привлекаются, когда требуются более высокие силы и вовлекают более крупные двигательные нейроны. У них обычно более короткое время сжатия, чем у меньших единиц. Количество дополнительных моторных единиц, задействованных во время данного приращения силы, резко снижается при высоких уровнях произвольной силы. Это говорит о том, что, хотя отряды с высоким порогом создают большее напряжение, вклад вербовки в увеличение произвольной силы снижается при более высоких уровнях силы.
Чтобы проверить стимуляцию двигательных единиц, электроды помещают внеклеточно на кожу и применяют внутримышечную стимуляцию. После стимуляции двигательной единицы ее импульс затем регистрируется электродом и отображается как потенциал действия , известный как потенциал действия двигательной единицы (MUAP). Когда несколько MUAP записываются в течение короткого промежутка времени, тогда отмечается поезд потенциала действия двигательной единицы (MUAPT). Время между этими импульсами известно как межимпульсный интервал (IPI). В медицинском электродиагностическом тестировании пациента со слабостью., тщательный анализ размера, формы и модели набора MUAP может помочь отличить миопатию от нейропатии .
Сокращение моторных единиц
Сокращение одной нейромоторной единицы зависит от ее функционального состояния, а целой мышцы — от количества функционирующих нейромоторных единиц. Наибольшее напряжение развивает нейромоторная единица икроножной и камбаловидной мышц, поддерживающих позу стояния.
При увеличении силы раздражения скелетной мышцы высота ее сокращения возрастает. Это зависит от количества возбужденных моторных единиц, число которых увеличивается по мере повышения силы раздражения (К. Люкас, 1910). Мышечные волокна, составляющие одну моторную единицу, сокращаются синхронно, одновременно, но мышечные волокна разных моторных единиц сокращаются, как правило, асинхронно, разновременно, так как разные моторные единицы иннервируются различными нейронами спинного мозга. В результате суммации сокращений отдельных моторных единиц получается гладкий тетанус, высота которого градуально повышается по мере увеличения числа сокращающихся моторных единиц. Следовательно, возрастание силы сокращения скелетной мышцы градуально зависит от числа сокращенных моторных единиц, а также обусловлено критическим пределом силы раздражения.
Сила сокращения мышцы зависит также от частоты возбуждения каждой моторной единицы. При небольшом напряжении мышцы частота импульсов возбуждения каждой моторной единицы равна 5-10 в 1 с, а при повышении напряжения — 20-50 до 150 в 1 с. Таким образом, высота сокращения мышцы возрастает также в зависимости ог частоты ее раздражения, но до известного критического предела.
Однако сила и частота раздражения определяют уровень обмена вещества в моторных единицах, который имеет решающее значение для градуального возрастания силы, или напряжения, скелетных мышц. Частота и сила импульсов регулируются по двигательным и вегетативным нервам скелетных мышц.
Сокращение и напряжение целой мышцы может долго продолжаться без утомления, так как в естественных условиях сокращение и напряжение мышц — результат суммации неодновременных, асинхронных сокращений и напряжений разных нейромоторных единиц.
Внешние ссылки [ править ]
| vтеМышечная ткань | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Гладкая мышца |
|
||||||||||||||||||||||||||||||||
| Поперечно-полосатая мышца |
|
Типы моторных единиц
Моторные единицы одной мышцы могут быть разными. В зависимости от скорости сокращения моторные единицы разделяют на медленные/slow (S-МЕ) и быстрые/fast (F-МЕ). А F-МЕ в свою очередь делят по устойчивости к утомлению на быстроутомляемые/fast-fatigable (FF-МЕ) и устойчивые к утомлению/fast-fatigue-resistant (FR-МЕ).
Соответствующим образом подразделяют иннервирующие данные МЕ мотонейроны. Существуют S-мотонейроны(S-МН), FF-мотонейроны (F-МН) и FR -мотонейроны(FR-МН). S- МЕ характеризуются высоким содержанием белка миоглобина, который способен связывать кислород (О2). Мышцы, преимущественно состоящие из МЕ этого типа, за их темно-красный цвет называются красными. Красные мышцы выполняют функцию поддержания позы человека. Предельное утомление таких мышц наступает очень медленно, а восстановление функций происходит наоборот, очень быстро.
Такая способность обуславливается наличием миоглобина и большого числа митохондрий. МЕ красных мышц, как правило, содержат большое количество мышечных волокон. FR-МЕ составляют мышцы, способные выполнять быстрые сокращения без заметного утомления. Волокна FR-ME содержат большое количество митохондрий и способны образовывать АТФ путём окислительного фосфорилирования.
Как правило, число волокон в FF-ME меньше, чем в S-ME. Волокна FR-ME характеризуются меньшим содержанием митохондрий, чем в FF-ME, а также тем, что АТФ в них образуется за счет гликолиза. В них отсутствует миоглобин, поэтому мышцы, состоящие из МЕ этого типа, называют белыми. Белые мышцы развивают сильное и быстрое сокращение, но довольно быстро утомляются.
Типы двигательных единиц (позвоночные) [ править ]
Двигательные единицы обычно классифицируются на основе сходства между несколькими факторами:
-
Физиологический
- Скорость сокращения при изометрических сокращениях
- Скорость нарастания силы
- Время до пика сокращения подергивания (ответ на одиночный нервный импульс)
- Скорость сокращения при изометрических сокращениях
-
- FF — Быстрая утомляемость — высокая сила, быстрая скорость сокращения, но утомляемость за несколько секунд.
- FR — Устойчивость к быстрой усталости — средняя сила, устойчивость к усталости — высокая скорость сжатия и устойчивость к усталости.
- FI — Fast Intermediate — промежуточное звено между FF и FR.
- S или SO — Медленный (окислительный) — низкая сила, меньшая скорость сокращения, высокая устойчивость к усталости.
-
Биохимический
- Гистохимический (самая старая форма биохимического типирования волокон)
- Активность гликолитического фермента (например, глицерофосфатдегидрогеназа (GPD) )
- Активность окислительного фермента (например, сукцинатдегидрогеназа -SDH)
- Чувствительность миозин- АТФазы к кислоте и щелочам
- Гистохимический (самая старая форма биохимического типирования волокон)
-
- Обычно они обозначают волокна как:
- I (медленное окисление, SO) — низкое гликолитическое и высокое окислительное присутствие. Низкая (er) миозиновая АТФаза, чувствительная к щелочам.
- IIa (Быстрый окислительный / гликолитический, FOG) — Высокая гликолитическая, окислительная и миозиновая АТФаза, чувствительная к кислоте.
- IIb (Fast glycolytic, FG) — высокий гликолитик и присутствие миозиновой АТФазы, чувствительный к кислоте. Низкое окислительное присутствие.
- IIi — волокна промежуточные между IIa и IIb.
-
- Гистохимический и физиологический типы соответствуют следующим образом:
- S и тип I, FR и тип IIa, FF и тип IIb, FI и IIi.
-
- Иммуногистохимический анализ (более поздняя форма типирования волокон)
- Тяжелая цепь миозина (MHC)
- Легкая цепь миозина — щелочь (MLC1)
- Легкая цепь миозина — регуляторная (MLC2)
- Иммуногистохимический анализ (более поздняя форма типирования волокон)
-
- Иммуногистохимические типы следующие, с типом IIa, IIb и медленным, соответствующим IIa, IIb и медленным (тип I) гистохимическим типам:
-
-
Выражено в Семья Джин Разработка Быстрые волокна (II) Медленные волокна (I) MHC Эмбриональный MHC MHC IIa β / медленный MHC Неонатальный MHC MHC IIb MHC IIx MLC1 (щелочь) Эмбриональный 1f 1 с 1f 3f MLC2 (нормативный) 2f 2f 2 с Таблица воспроизведена из
-
-
- В настоящее время известно около 15 различных типов генов MHC, распознаваемых в мышцах, только некоторые из них могут экспрессироваться в одном мышечном волокне. Эти гены образуют один из ~ 18 классов генов миозина, идентифицированных как класс II, который не следует путать с миозинами типа II, идентифицированными с помощью иммуногистохимии. Экспрессия нескольких генов MHC в одном мышечном волокне является примером полиморфизма . Относительная экспрессия этих типов миозина определяется частично генетикой и частично другими биологическими факторами, такими как активность, иннервация и гормоны.
Таким образом, типизация двигательных единиц прошла через много этапов и достигла точки, когда было признано, что мышечные волокна содержат различные смеси нескольких типов миозина, которые не могут быть легко разделены на определенные группы волокон. Три (или четыре) классических типа волокон представляют собой пики в распределении свойств мышечных волокон, каждый из которых определяется общей биохимией волокон.
Оценки коэффициентов иннервации двигательных единиц в мышцах человека:
| Мышцы | Количество моторных аксонов | Количество мышечных волокон | Коэффициент иннервации | Справка |
|---|---|---|---|---|
| Бицепс | 774 | 580 000 | 750 | Бухталь, 1961 г. |
| Брахиорадиалис | 315 | 129 000 | 410 | Файнштейн и др. |
| Первый спинной межкостный | 119 | 40 500 | 340 | Файнштейн и др. |
| Медиальная икроножная мышца | 579 | 1,120,000 | 1,934 | Файнштейн и др. |
| Передняя большеберцовая мышца | 445 | 250 200 | 562 | Файнштейн и др. |
Таблица воспроизведена из Карпати (2010)
«Принцип величины» Хеннемана
Увеличение нагрузки вызывает возбуждение различных типов МН в соответствии с их размерами. Порядок вовлечение новых МН, таким образом, как правило, одинаков практически при любом виде сокращения: сначала в процесс вовлекаются МН меньшего размера, затем большего. Такую закономерность в 1956 году Эдвуд Хеннеман описал как «принцип величины».
Еще до Хенемана ряд ученых описали некоторые положения этой закономерности. В частности, Денни-Броун и Эдриан Бронк, году исследовали принципы работы мышечной единицы. В 1929 они предположили, что есть два способа, с помощью которых нервная система может управлять сокращением мышц:
- увеличение частоты подаваемых импульсов
- увеличение количества вовлеченных в процесс сокращения МН.
В 1938 году Денни-Броун и Пеннибекер привели основные положения принципа величины в отношении МЕ, в отличие от Хеннемана, который сначала говорил только о МН, связывая порядок их вовлечения в работу с их размерами.
Мотонейронный пул
Мотонейроны, иннервирующие одну мышцу, составляют общий мотонейронный пул. В одном мотонейронном пуле могут находиться мотонейроны разных размеров. Крупные мотонейроны, имеющие толстые аксоны и множество коллатералей, взаимодействующие с большим числом мышечных волокон содержатся в крупных МЕ. Такие МЕ характеризуются высокой скоростью проведения возбуждения, при этом имеют низкую возбудимость и генерируют высокую частоту нервных импульсов (20—50 имп./с).
Более мелкие МЕ включают в себя МН небольших размеров, имеющие медленнопроводящие тонкие аксоны и взаимодействующие с небольшим числом мышечных волокон. Крупные МН возбуждается только при больших нагрузках на мышцу, а мелкие МН могут включаться в работу при небольших мышечных усилиях.
Нарушения «принципа величины»
Изначально предполагалось, что принцип величины работает при увеличивающемся изометрическом сокращении. Изометрическое сокращение — это сокращение мышцы без изменения её длины. Мышца сокращается изометрически при совершении статической работы. (Например, вы упираетесь плечом в стену, но сдвинуть её не можете.) Но мышечное сокращение не всегда является изометрическим (есть ещё изотоническое и ауксоническое). Даже в тех мышцах, на примере которых можно изучать изометрическое сокращение, одна и та же моторная единица может иметь разные пороги для активации (вовлечения) следующего своего звена для сгибания и разгибания. Такие замечания, которые часто обозначаются как «задания специфического ответа мотонейронов» (Ericksson et al., 1984), описали действие жевательного мускула человека (English, 1985) и подтвердили, что не все моторные единицы задействованы во время мышечного сокращения.
Идея о том, что существуют отдельные группы мотонейронов, отвечающие по-разному на какое-либо направленное движение, является исключением из «принципа величины» как для мышц челюсти, так и для мышц конечностей. До некоторого времени предполагалось что в зависимости от типа движения включается одна или другая моторная единица, однако позже было доказано, что это на самом деле две мышцы с двумя мотонейронными пулами, то есть эта мышца находится в процессе разделения на две различные. Весьма убедительный пример был приведён новозеландским неврологом Дереком Денни-Броуном, который в 1949 году показал, что при «хватательном» движении кисти в мышце flexor profundus digitorum моторные единицы включаются в одном порядке, а при «сгибательном» движении — в другом порядке.
Так же был проведён ряд экспериментов на интактных и децеребрированных кошках. Сначала в 1970 году группа ученых (Берк, Янковская, Тен Бруггенкате) в ходе своих исследований обнаружили что FF и FR мотонейроны могут быть полисинаптически возбуждены входами от низкопороговых кожных афферентов, а S-мотонейроны наоборот могут быть теми же входами заторможены. При этом возбуждающий постсинаптический потенциал (ВПСП) на F-мотонейронах является дисинаптическим, а на S-мотонейронах трисинаптическим (Иллерт и др. 1976). В 1982 году было показано что этот эффект вызывается моторной корой и красным ядром (см. Головной мозг) (Burke, 1982)
Рефлекс «отряхивания лапы»
Затем был открыт рефлекс, являющийся примером ситуации, когда необходимо включить только быстрые моторные единицы, без использования медленных. Это натуральный спинальный рефлекс «встряхивания лапы». Этот рефлекс сохраняется у спинальных кошек, и естественно присутствует у особей с интактной нервной системой. «Встряхивание» наблюдается у интактной кошки, когда та наступит лапой в воду. Следовательно, за запуск данного рефлекса отвечают низкопороговые кожные афференты подушечек лапы. В этом рефлексе не задействован soleus («медленная» мышца голени см. Нога, время сокращения 20-25 мс), а работает только gastrocnemius («быстрая» мышца голени, время сокращения 80 мс). Частота отряхивания лапы очень велика, настолько (10-12Гц), что исключает возможность задействования медленных моторных единиц.
В 1980 году Д.Смит (Д.Смит и др. Дж. физиол. 1980) в своей работе «Быстрые экстензоры голеностопа во время встряхивания лапы: их избирательное вовлечение» описал изучение солиуса и гастрокнемиуса кошки в трёх состояниях (стояние, ходьба, прыжки). Оказалось, что обе эти мышцы работают вместе во всех трёх случаях. То же было показано для быстрых и медленных мышц-разгибателей передних конечностей. Оказалось, что несмотря на то, что солиус является медленной мышцей, это нисколько не мешает при быстрых движениях (галоп, прыжки). И опять же Смит и др. попытались найти такое движение, которое было бы слишком быстрым для солиуса. Солиус способен развить усилие за 80 мс (время разгибания сустава при прыжке на 1 м = 130—150 мс). Смит также обнаружил что частота встряхивания лапы близка по частоте к тремору (10-13 раз в сек), следовательно, при таком быстром движении солиус молчит. Однако, если частота чесания приближается к 120—150 мс солиус работает (!), как и при прыжке.
В 1999 г. учёными из Атланты (Коп и Соколов) было доказано, что для медиальной и латеральной головок гастрокнемиуса «принцип величины Хеннемана» вполне применим и значительно облегчает координацию работы различных мышц. При этом они исследовали применения этого принципа к пулам мотонейронов мышц, коактивирующихся (такое движение, при котором мышца-антагонист или группа мышц всё же частично активны, когда начальная мышечная активность исчерпалась. Например, трицепс частично активен, когда бицепс совершает какие-либо маневры, а квадрицепс частично активизирован, когда активен гамстринг) в этом конкретном движении.






