Потенциал покоя и потенциал действия. мембранный потенциал покоя это разность электрических потенциалов (напряжение) между внутренней и наружной поверхностью. — презентация
Содержание:
- Введение
- ПОТЕНЦИАЛ ПОКОЯ МЕМБРАНЫ МЫШЕЧНОГО ВОЛОКНА И ПОТЕНЦИАЛ ДЕЙСТВИЯ
- Физиологические свойства возбудимых тканей
- Потенциалы покоя
- Определение
- Возбудимость
- Физиология формирования мембранного потенциала действия
- Уравнения Голдмана
- Электронейтральность
- Уравнение Нернста
- Формирование потенциала покоя
- Равновесный потенциал
- Состав плазматической мембраны
Введение
Зачем нам нужно знать, что такое потенциал покоя?
Что такое «животное электричество»? Откуда в организме берутся «биотоки»? Как живая клетка, находящаяся в водной среде, может превратиться в «электрическую батарейку»?
— На эти вопросы мы сможем ответить, если узнаем, как клетка за счёт перераспределения электрических зарядов создаёт себе электрический потенциал на мембране.
Как работает нервная система? С чего в ней всё начинается? Откуда в ней берётся электричество для нервных импульсов?
— На эти вопросы мы также сможем ответить, если узнаем, как нервная клетка создаёт себе электрический потенциал на мембране.
Итак, понимание того, как работает нервная система, начинается с того, что надо разобраться, как работает отдельная нервная клетка — нейрон.
А в основе работы нейрона с нервными импульсами лежит перераспределение электрических зарядов на его мембране и изменение величины электрических потенциалов. Но чтобы потенциал изменять, его нужно для начала иметь. Поэтому можно сказать, что нейрон, готовясь к cвоей нервной работе, создаёт на своей мембране электрический потенциал, как возможность для такой работы.
Таким образом, наш самый первый шаг к изучению работы нервной системы — это понять, каким образом перемещаются электрические заряды на нервных клетках к как за счёт этого на мембране появляется электрический потенцила. Этим мы и займёмся, и назовём этот процесс появления электрического потенциала у нейронов — формирование потенциала покоя.
ПОТЕНЦИАЛ ПОКОЯ МЕМБРАНЫ МЫШЕЧНОГО ВОЛОКНА И ПОТЕНЦИАЛ ДЕЙСТВИЯ
Давайте рассмотрим, как возникает и развивается потенциал действия, который приводит в дальнейшем к сокращению скелетных мышц. Вначале разберем, что такое потенциал покоя.
Потенциал покоя мембраны мышечного волокна
В состоянии покоя сарколемма (мембрана) мышечного волокна поляризована или, другими словами, имеется определенный мембранный потенциал покоя. Снаружи мембраны заряд положительный, а внутри – отрицательный (рис.1). Разность потенциалов между наружной и внутренней оболочками мембраны мышечного волокна составляет 90 мВ.
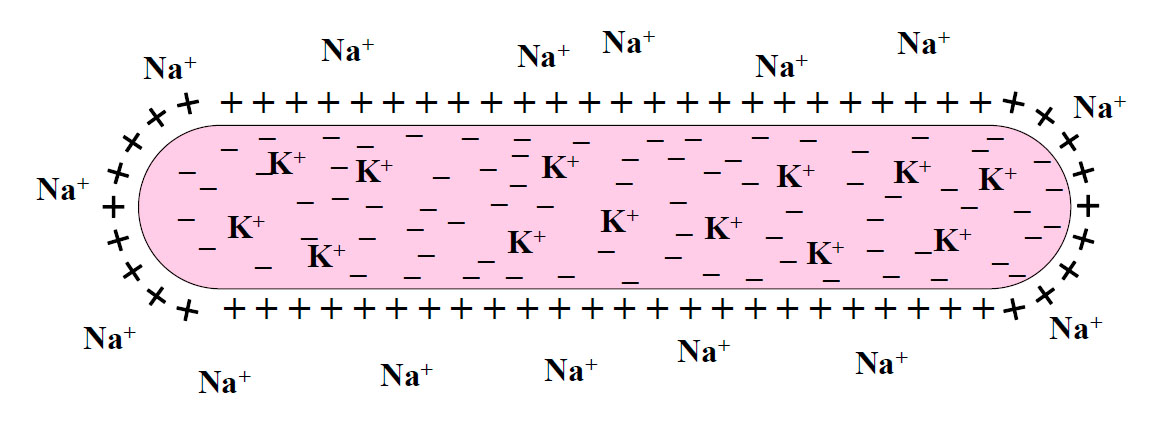
Рис.1
В тканевой жидкости, окружающей мышечные волокна, выше концентрация ионов натрия (Na+), а в саркоплазме мышечного волокна – ионов калия (К+). Однако положительно заряженные ионы К+ не полностью уравновешивают анионы (отрицательно заряженные ионы), содержащиеся в саркоплазме мышечного волокна, это обусловливает отрицательный заряд мембраны мышечного волокна (то есть ее внутренней оболочки).
Более подробно строение и функции мышц описаны в моих книгах:
- Гипертрофия скелетных мышц человека
- Биомеханика опорно-двигательного аппарата человека
мышечное волокнамотонейронамышечного волокнамышечное волокно++мышечное волокно+мышечного волокнадеполяризуется
Потенциал действия
Возникшая волна деполяризации передается вдоль оболочки мышечного волокна. При этом все больше открывается каналов натрия и все больше ионов Na+ входит внутрь волокна. Скорость проникновения ионов Na+ внутрь мышечного волокна очень высокая — несколько миллионов ионов в секунду (А. Дж. Мак-Комас, 2001) (рис.2).
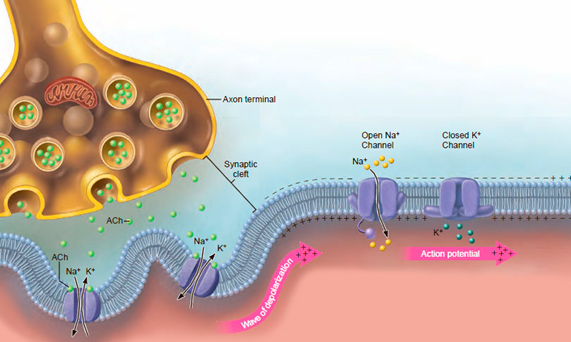
Рис. 2. Распространение волны деполяризации вдоль поверхностной мембраны мышечного волокна (E.N.Marieb, 2015)
Каналы калия, однако остаются закрытыми. Через каналы натрия ионы К+ пройти не могут. Это связано с тем, что ионы Na+ имеют диаметр 0,1 нм, а ионы К+ — 0,13 нм.
Этот кратковременный процесс (не более 1-2 мс) деполяризации мышечного волокна называется потенциалом действия. Разность потенциалов между оболочками мышечного волокна доходит до 120-130 мВ. Волна деполяризации через Т-трубочки достигает саркоплазматического ретикулума, и из него в саркоплазму выделяются ионы кальция (Ca2+) начинается процесс сокращения мышечного волокна. Об этом я расскажу более подробно в дальнейшем.
Следует заметить, что процесс распространения волны деполяризации вдоль мышечного волокна можно зарегистрировать посредством электромиографии.
Реполяризация
После прохождения волны деполяризации, каналы натрия закрываются и открываются каналы калия. Ионы К+ начинают выходить из мышечного волокна, так как они заряжены положительно, а снаружи мембрана заряжена отрицательно. Потенциал действия снижается. Мембрана мышечного волокна восстанавливает свою полярность. Это называется реполяризацией. Вновь снаружи она заряжена положительно, а внутри – отрицательно. Однако существуют отличия от первоначального состояния мышечного волокна, так как снаружи мышечного волокна теперь много ионов К+, а внутри мышечного волокна много ионов Na+ .
Работа натрий-калиевой помпы (насоса)
Чтобы восстановить исходное состояние мышечного волокна начинает действовать натрий-калиевый насос (помпа). Этот насос за счет энергии АТФ активно выкачивает из мышечного волокна ионы Na+ и закачивает ионы К+ внутрь. Натрий-калиевый насос представляет собой белковую молекулу. Таких молекул в мембране мышечного волокна достаточно много. На работу этого механизма тратится около 70% энергии мышечного волокна.
Работа кальциевой помпы (насоса)
Чтобы закачать в саркоплазматический ретикулум ионы кальция, начинает работать кальциевый насос. Этот насос закачивает в саркоплазматический ретикулум 90% ионов кальция (Ca2+). Функционирование этого насоса стимулирует присутствие ионов магния ( Mg2+). Для транспорта ионов кальция в саркоплазматический ретикулум также нужна энергия АТФ. Доказано, что для транспорта двух ионов кальция тратится одна молекула АТФ ( А. Дж. МакКомас, 2001).
Литература:
1. Мак-Комас А. Дж. Скелетные мышцы человека. – Киев: Олимпийская литература, 2001.- 407 с.
Физиологические свойства возбудимых тканей
Основными свойствами всех возбудимых тканей являются:
- возбудимость,
- проводимость,
- лабильность.
Лабильность (функциональная подвижность) — это способность ткани реагировать на различные воздействия с определенной скоростью, то есть способность воспроизводить определенную частоту раздражения.
Мерой лабильности является наибольшее число ответных реакций, которыми возбудимая ткань способна реагировать за единицу времени в соответствии с частотой приложенных к ней раздражений.
Проводимость — это способность ткани и клетки передавать возбуждение.
Раздражение — это процесс воздействия на живую ткань агентов внешней по отношению к этой ткани среды.
Раздражитель — это причина, способная вызвать ответную реакцию.
Потенциалы покоя
Мембранный потенциал покоя не является равновесным потенциалом, так как он зависит от постоянного расхода энергии (для ионных насосов, как упоминалось выше) на его поддержание. Этот механизм учитывается динамическим диффузионным потенциалом — полностью в отличие от равновесного потенциала, который справедлив независимо от природы рассматриваемой системы. В мембранном потенциале покоя доминируют ионные частицы в системе, которые имеют наибольшую проводимость через мембрану. Для большинства клеток это калий. Поскольку калий также является ионом с наиболее отрицательным равновесным потенциалом, обычно потенциал покоя может быть не более отрицательным, чем равновесный потенциал калия. Потенциал покоя может быть рассчитан с помощью уравнения напряжения Гольдмана-Ходжкина-Каца с использованием концентраций ионов как для равновесного потенциала, а также с учетом относительной проницаемости каждого вида ионов . В нормальных условиях можно с уверенностью предположить, что только ионы калия, натрия (Na + ) и хлорида (Cl — ) играют большую роль в потенциале покоя:
- E м знак равно р Т F пер ( п N а + N а + о + п K + K + о + п C л — C л — я п N а + N а + я + п K + K + я + п C л — C л — о ) {\ displaystyle E_ {m} = {\ frac {RT} {F}} \ ln {\ left ({\ frac {P_ {Na ^ {+}} _ {o} + P_ { K ^ {+}} _ {o} + P_ {Cl ^ {-}} _ {i}} {P_ {Na ^ {+}} _ {i} + P_ {K ^ {+}} _ {i} + P_ {Cl ^ {-}} _ {o}}} \верно)}}
Это уравнение напоминает уравнение Нернста, но имеет член для каждого проникающего иона. Кроме того, в уравнение был вставлен z , в результате чего внутриклеточные и внеклеточные концентрации Cl — меняются местами относительно K + и Na + , поскольку отрицательный заряд хлорида обрабатывается путем инвертирования дроби внутри логарифмического члена. * E m — мембранный потенциал, измеренный в вольтах * R , T и F такие, как указано выше * P s — относительная проницаемость иона s * Y — концентрация иона s в отсеке Y, как указано выше. Другой способ увидеть мембранный потенциал, учитывая вместо этого проводимость ионных каналов, а не проницаемость мембраны, — это использовать уравнение Миллмана (также называемое уравнением хордовой проводимости):
- E м знак равно грамм K + E е q , K + + грамм N а + E е q , N а + + грамм C л — E е q , C л — грамм K + + грамм N а + + грамм C л — {\ displaystyle E_ {m} = {\ frac {g_ {K ^ {+}} E_ {eq, K ^ {+}} + g_ {Na ^ {+}} E_ {eq, Na ^ {+}} + g_ {Cl ^ {-}} E_ {eq, Cl ^ {-}}} {g_ {K ^ {+}} + g_ {Na ^ {+}} + g_ {Cl ^ {-}}}}}
или переформулирован
- E м знак равно грамм K + грамм т о т E е q , K + + грамм N а + грамм т о т E е q , N а + + грамм C л — грамм т о т E е q , C л — {\ displaystyle E_ {m} = {\ frac {g_ {K ^ {+}}} {g_ {tot}}} E_ {eq, K ^ {+}} + {\ frac {g_ {Na ^ {+}) }} {g_ {tot}}} E_ {eq, Na ^ {+}} + {\ frac {g_ {Cl ^ {-}}} {g_ {tot}}} E_ {eq, Cl ^ {-}} }
где g tot — общая проводимость всех ионных частиц, снова в произвольных единицах. Последнее уравнение изображает мембранный потенциал покоя как средневзвешенное значение реверсивных потенциалов системы, где веса представляют собой относительные проводимости каждого вида ионов ( г X / г итого ). Во время потенциала действия эти веса изменяются. Если проводимости Na + и Cl — равны нулю, мембранный потенциал уменьшается до потенциала Нернста для K + (как g K + = g tot ). Обычно в условиях покоя g Na + и g Cl− не равны нулю, но они намного меньше, чем g K + , что делает E m близким к E eq, K + . Медицинские условия , такие как гиперкалиемия , в которых в крови в сыворотке калия (который регулирует O ) изменяются очень опасны , так как они смещены E э, K + , влияя таким образом на E м . Это может вызвать аритмию и остановку сердца . Использование болюсной инъекции хлорида калия при казнях путем останавливает сердце, сдвигая потенциал покоя к более положительному значению, что деполяризует и сокращает сердечные клетки навсегда, не позволяя сердцу реполяризоваться и, таким образом, входить в диастолу для пополнения с кровью.
Хотя уравнение напряжения GHK и уравнение Миллмана связаны, они не эквивалентны. Критическое различие состоит в том, что уравнение Миллмана предполагает, что отношение тока к напряжению является омическим, тогда как уравнение напряжения GHK учитывает небольшие мгновенные выпрямления, предсказываемые уравнением потока GHK, вызванные градиентом концентрации ионов. Таким образом, более точная оценка мембранного потенциала может быть рассчитана с использованием уравнения GHK, чем с помощью уравнения Миллмана.
Определение
В норме, когда клетка готова к работе, у неё уже есть электрический заряд на поверхности мембраны. Он называется мембранный потенциал покоя.
Потенциал покоя — это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. Его средняя величина составляет -70 мВ (милливольт).
«Потенциал» — это возможность, он сродни понятию «потенция». Электрический потенциал мембраны — это её возможности по перемещению электрических зарядов, положительных или отрицательных. В роли зарядов выступают заряженные химические частицы — ионы натрия и калия, а также кальция и хлора. Из них только ионы хлора заряжены отрицательно (-), а остальные — положительно (+).
Таким образом, имея электрический потенциал, мембрана может перемещать в клетку или из клетки указанные выше заряженные ионы.
Важно понимать, что в нервной системе электрические заряды создаются не электронами, как в металлических проводах, а ионами — химическими частицами, имеющими электрический заряд. Электрический ток в организме и его клетках — это поток ионов, а не электронов, как в проводах
Обратите также внимание на то, что заряд мембраны измеряется изнутри клетки, а не снаружи
Если говорить уж совсем примитивно просто, то получается, что снаружи вокруг клетки будут преобладать «плюсики», т.е. положительно заряженные ионы, а внутри — «минусики», т.е. отрицательно заряженные ионы. Можно сказать, что внутри клетка электроотрицательна. И теперь нам всего лишь надо объяснить, как это так получилось. Хотя, конечно, неприятно сознавать, что все наши клетки — отрицательные «персонажи». ((
Сущность
Сущность потенциала покоя — это преобладание на внутренней стороне мембраны отрицательных электрических зарядов в виде анионов и недостаток положительных электрических зарядов в виде катионов, которые сосредотачиваются на её наружной стороне, а не на внутренней.
Внутри клетки — «отрицательность», а снаружи — «положительность».
Такое положение вещей достигается с помощью трёх явлений: (1) поведения мембраны , (2) поведения положительных ионов калия и натрия и (3) соотношения химической и электрической силы.
1. Поведение мембраны
В поведении мембраны для потенциала покоя важны три процесса:
1) Обмен внутренних ионов натрия на наружные ионы калия. Обменом занимаются специальные транспортные структуры мембраны: . Таким способом мембрана перенасыщает клетку калием, но обедняет натрием.
2) Открытые калиевые ионные каналы. Через них калий может как заходить в клетку, так и выходить из неё. Он выходит в основном.
3) Закрытые натриевые ионные каналы. Из-за этого натрий, выведенный из клетки насосми-обменниками, не может вернуться в неё обратно. Натриевые каналы открываются только при особых условиях — и тогда потенциал покоя нарушается и смещается в сторону нуля (это называется деполяризацией мембраны, т.е. уменьшением полярности).
2. Поведение ионов калия и натрия
Ионы калия и натрия по-разному перемещаются через мембрану:
1) Через ионные насосы-обменники натрий насильно выводится из клетки, а калий затаскивается в клетку.
2) Через постоянно открытые калиевые каналы калий выходит из клетки, но может и возвращаться в неё обратно через них же.
3) Натрий «хочет» войти в клетку, но «не может», т.к. каналы для него закрыты.
3. Соотношение химической и электрической силы
По отношению к ионам калия между химической и электрической силой устанавливается равновесие на уровне — 70 мВ.
1) Химическая сила выталкивает калий из клетки, но стремится затянуть в неё натрий.
2) Электрическая сила стремится затянуть в клетку положительно заряженные ионы (как натрий, так и калий).
Возбудимость
Возбудимость — способность ткани отвечать на раздражение специализированной реакцией — возбуждением. Возбудимость является одной из форм раздражимости.
Возбудимыми тканями называются такие ткани, клетки которых способны в ответ на раздражение генерировать специфическую реакцию — возбуждение.
Возбуждение — это специализированная ответная реакция живого объекта на действие раздражителя, проявляющаяся в изменениях его определенных параметров.
К возбудимым тканям относятся:
- нервная,
- мышечная,
- железистая.
Признаки возбуждения:
- общие,
- специфические.
Общие признаки возбуждения (присущи всем возбудимым тканям):
- Изменение уровня обменных процессов в тканях;
- Выделение различных видов энергии — тепловой, электрической.
Специфические признаки возбуждения (характерны для определенного вида тканей):
- Мышечная ткань — сокращение,
- Железистая — выделение секрета,
- Нервная — генерация и проведение нервного импульса.
Невозбудимыми являются:
- эпителиальная,
- соединительная ткани.
Физиология формирования мембранного потенциала действия
Потенциал действия — это быстрый рост и последующее падение напряжения или мембранного потенциала через клеточную мембрану с характерным рисунком. Достаточный ток необходим для инициирования отклика напряжения в клеточной мембране; если ток недостаточен для деполяризации мембраны до порогового уровня, потенциал действия не будет срабатывать. Примерами клеток, передающих сигналы через потенциалы действия, являются нейроны и мышечные клетки.
Стимул запускает быстрое изменение напряжения или потенциала действия. В режиме патч-зажим в ячейку должен подаваться достаточный ток, чтобы поднять напряжение выше порогового напряжения, чтобы начать деполяризацию мембраны.
Деполяризация вызвана быстрым увеличением мембранного потенциала , открытием натриевых каналов в клеточной мембране, что приводит к большому притоку ионов натрия.
Реполяризация мембраны является результатом быстрой инактивации натриевых каналов, а также большого оттока ионов калия в результате активированных калиевых каналов.
Гиперполяризация — это пониженный мембранный потенциал , вызванный истечением ионов калия и закрытием калиевых каналов. Состояние покоя — это когда мембранный потенциал возвращается к напряжению покоя, которое возникло до того, как возник стимул. У вашего тела есть нервы, которые соединяют ваш мозг с остальными органами и мышцами, точно так же, как телефонные провода соединяют дома по всему миру. Когда вы хотите, чтобы ваша рука двигалась, ваш мозг посылает сигналы через нервы к вашей руке, сообщая мышцам сокращаться. Но ваши нервы не просто говорят «рука, двигайтесь». Вместо этого ваши нервы посылают множество электрических импульсов (так называемых потенциалов действия) различным мышцам в вашей руке, что позволяет вам двигаться с предельной точностью.
Нейроны представляют собой особый тип клеток с единственной целью передачи информации по всему телу. Нейроны похожи на другие клетки тем, что имеют клеточное тело с ядром и органеллами. Тем не менее, у них есть несколько дополнительных функций, которые позволяют им быть фантастическими при передаче потенциалов действия:
- дендриты: получают сигналы от соседних нейронов (например, радиоантенны)
- аксон: передавать сигналы на расстоянии (например, телефонные провода)
- терминал аксона: передавать сигналы другим дендритам или тканям нейронов (например, радиопередатчик)
- миелиновая оболочка: ускоряет передачу сигнала по аксону
Уравнения Голдмана
Хотя равновесный потенциал для ионов калия составляет -90 мВ, МПС нейрона несколько менее отрицательный. Эта разница отражает незначительное но постоянное следование ионов Na + через мембрану в состоянии покоя. Поскольку концентрационный градиент для натрия противоположный такого для калия, Na + движется внутрь клетки и сдвигает суммарный заряд на внутренней стороне мембраны в положительную сторону. На самом деле МПС нейрона составляет от -60 до -80 мВ. Это значение значительно ближе к Е K чем до Е Na, потому что в состоянии покоя в нейроне открыто много калиевых каналов и очень мало натриевых. Также на встанвлення МПС влияет движение ионов хлора. В 1943 году Дэвид Голдаман предложил усовершенствовать уравнение Нернста так, чтобы оно отражало влияние различных ионов на мембарнний потенциал, в этом уравнении учитывается относительная проницаемость мембраны для каждого типа ионов:
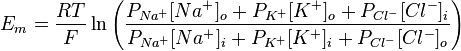
где R — универсальная газовая постоянная, Т — абслоютна температура (по шкале Кельвина), z — заряд иона, F — число Фарадея, o,i — концентрации ионов внутри и внутри клеток, Р — относительная проницаемость мембраны для соответствующего иона. Значение заряда в данном уравнении не сохраняется, но оно учтено — для хлора внешняя и внутренняя концентрация поменяны местами, так как его заряд 1.
Электронейтральность
В большинстве количественных обработок мембранного потенциала, таких как вывод уравнения Гольдмана , предполагается электронейтральность ; это означает, что на любой стороне мембраны нет измеримого превышения заряда. Таким образом, хотя существует электрический потенциал на мембране из-за разделения зарядов, нет реально измеримой разницы в общей концентрации положительных и отрицательных ионов на мембране (как это оценивается ), то есть нет реально измеримых превышение заряда с обеих сторон. Это происходит потому, что влияние заряда на электрохимический потенциал намного больше, чем эффект концентрации, поэтому необнаружимое изменение концентрации вызывает большое изменение электрического потенциала.
Уравнение Нернста
Равновесный потенциал для определенного иона, например для калия, можно рассчитать по уравнению Нернста, что выглядит так:
- ,
где R — универсальная газовая постоянная, Т — абслоютна температура (по шкале Кельвина), z — заряд иона, F — число Фарадея, o,i — концентрация калия снаружи и внутри клетки соответственно. Поскольку описанные процессы происходят при температуре тела — 310 ° К, а десятичными логарифмами в исчислении пользоваться легче чем натуральными, это уравнение превращают следующим образом:
Подставляя концентрации К + в уравнение Нернста получаем равновесный потенциал для калия, составляет -90 мВ. Поскольку по нулевой потенциал принимается внешняя сторона мембраны, то знак минус означает, что в условиях равновесного калиевого потенциала внутренняя Сторн мембраны сравнительно более электроотрицательным. Аналогичные расчеты можно провести и для равновесного Натиева потенциала, он составляет 62 мВ.
Формирование потенциала покоя
ПП формируется в два этапа.
Первый этап:
создание незначительной (-10 мВ) отрицательности внутри клетки за счёт неравного асимметричного обмена Na + на K + в соотношении 3: 2. В результате этого клетку покидает больше положительных зарядов с натрием, чем возвращается в неё с калием. Такая особенность работы натрий-калиевого насоса, осуществляющего взаимообмен этих ионов через мембрану с затратами энергии АТФ , обеспечивает его электрогенность.
Результаты деятельности мембранных ионных насосов-обменников на первом этапе формирования ПП таковы:
1. Дефицит ионов натрия (Na +) в клетке.
2. Избыток ионов калия (K +) в клетке.
3. Появление на мембране слабого электрического потенциала (-10 мВ).
Второй этап:
создание значительной (-60 мВ) отрицательности внутри клетки за счёт утечки из неё через мембрану ионов K + . Ионы калия K + покидают клетку и уносят с собой из неё положительные заряды, доводя отрицательность до −70 мВ.
Итак, мембранный потенциал покоя — это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса.
Равновесный потенциал
Для того, чтобы определить влияние движения определенного иона через полупроницаемую мембрану на формирование мембранного потенциала, строят модельные системы. Такая модельная система состоит из сосуда разделенной на две ячейки искусственной полупроницаемой мембраной, в которую встроены ионные каналы. В каждую ячейку можно погрузить электрод и померить разность потенциалов.
Рассмотрим случай, когда искусственная мембрана проницаема только для калия. По две стороны мембраны модельной системы создают градиент концентрации аналогичный таковому у нейроне: в ячейку, соответствующую цитоплазме (внутренняя ячейка), помещают 140 мМ раствор хлорида калия (KCl), в ячейку, соответствующую межклеточной жидкости (внешняя ячейка) — 5 ммоль раствор KCl. Ионы калия будут диффундировать через мембрану во внешнюю ячейку по градиенту концентрации. Но поскольку анионы Cl — проникать через мембрану не могут во внутренней ячейке возникать избыток отрицательного заряда, который будет препятствовать подали оттока катионов. Когда такие модельные нейроны достигнут состояния равновесия, действие химического и электрического потенциала будет сбалансирована, ни суммарной диффузии К + не будет наблюдаться. Значение мембранного потенциала, виинкае при таких условиях, называется равновесным потенциалом для определенного иона (Е ион). Равновесный потенциал для калия составляет примерно -90 мВ.
Аналогичный опыт можно провести и для натрия, установив между ячейками мембрану проникающей только для этого катиона, и поместив во внешнюю ячейку раствор хлорида натрия с концентрацией 150 мМ, а во внутреннюю — 15 мМ. Натрий будет двигаться во внутреннюю ячейку, ривоноважний потенциал для него составит примерно 62 мВ.
Количество ионов, должна диффундировать для генерации электрического потенциала очень невелика (примерно 10 -12 моль К + на 1 см 2 мембраны), этот факт имеет два важных последствия. Во-прешь, это означает, что концентрации ионов, которые могут проникать через мембрану, остаются стабильными снаружи и внутри клетки, даже после того как их движение обеспечил утоврення электрического потенциала. Во-вторых, мизерные потоки ионов через мембрану, потирбно для установления потенциала, не нарушают электронейтральности цитоплазмы и внеклеточной жидкости в целом, распределение зарядов происходит только в области, непосредственно прилегающей к плазматической мембраны.
Состав плазматической мембраны
- Липиды (в основном фосфолипиды),
- Белки (гликопротеины),
- Углеводы (мукополисахариды).
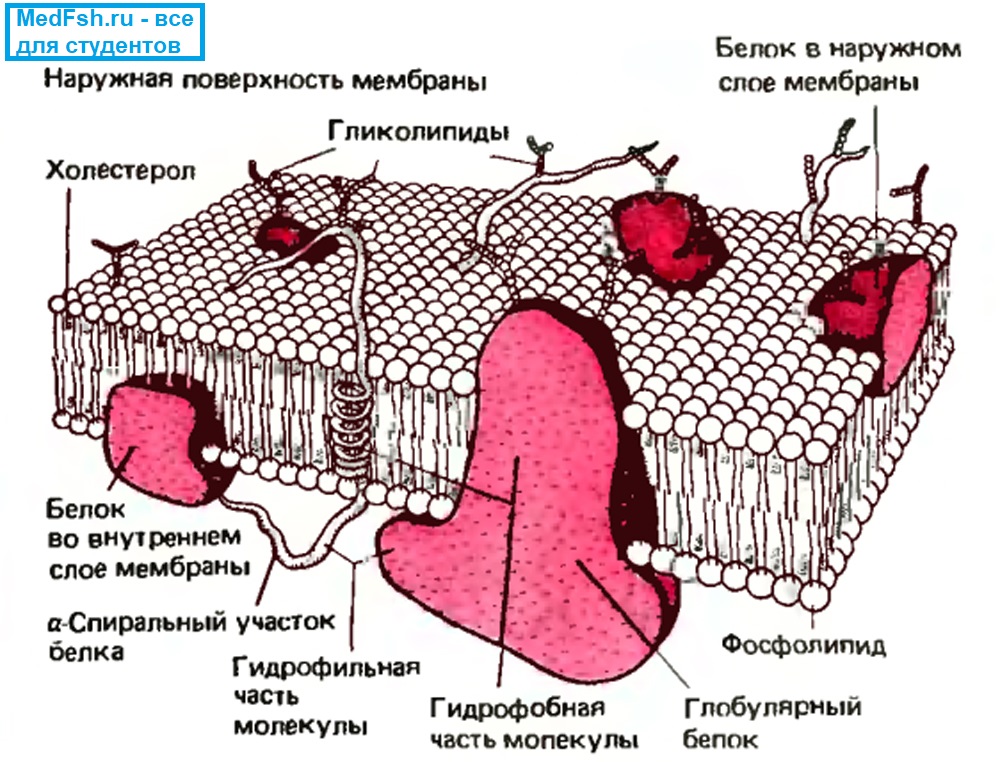
Липиды очень плотно упакованы в мембране, между ними нет практически никаких расстояний, поэтому мембрана плохо пропускает воду, практически непроницаема для ионов и других крупных молекул.
Белковые молекулы могут быть погружены в слой липидов с внеклеточной или с цитоплазматической стороны, либо могут целиком пронизывать мембрану.
Если белки прикреплены к поверхности мембраны, их называют периферическими. С внутренней стороны это будут белки-ферменты, а с наружной — белки-рецепторы.
Если белки пронизывают всю толщу мембраны клеток, то их называют интегральными или трансмембранными.
Такие белки образуют структуры, обеспечивающие движение ионов через мембрану.
Если белки образуют стенки поры, сквозь которую путем простой диффузии проходят ионы, то это ионные каналы.
Если трансмембранные белки перекачивают ионы против концентрационного и электрического градиентов, то это ионные насосы.
Все каналы, имеющиеся в живых тканях, можно разделить на 2 типа:
- первый тип — каналы покоя, которые спонтанно открываются и закрываются без всяких внешних воздействий;
- второй тип — gate-каналы (воротные каналы) — в покое они закрыты и открываются под действием раздражителей.
Ионные каналы:
- неспецифические (каналы утечки, всегда открыты),
- специфические (селективные), обладающие способностью пропускать только определенные ионы при изменении заряда на мембране или действии химических веществ.






