Анатомия рецепторов человека
Содержание:
- Введение
- Клеточные рецепторы
- Классификация рецепторов
- Виды некоторых рецепторов и их данные[править | править код]
- Рецепторы человека[править | править код]
- Виды некоторых рецепторов и их данные
- Лиганд (биохимия)
- Системы вторых посредников
- Антиноцицептивные системы и рецепторы
- Визуализация эффектов адреналина
- Выманивание рецепторов из потаенных мест
- Секретные PRR
Введение
Рецептор — воспринимающий элемент; чаще всего — в биологии. Это сложное образование, состоящие из терминалей (нервных окончаний) дендритов чувствительных нейронов, глии, специализированных образований межклеточного вещества и специализированных клеток других тканей, которые в комплексе обеспечивают превращение влияния факторов внешней или внутренней среды (раздражитель) в нервный импульс — волна возбуждения, которая распространяется по нервному волокну и служит для передачи информации от периферических рецепторных (чувствительных) окончаний к нервным центрам, внутри центральной нервной системы и от неё к исполнительным аппаратам — мышцам и железам. Они состоят из «терминалей», дендрита нейронов, глий, специализированных образований межклеточного вещества и специализированных клеток иных тканей. В некоторых рецепторах (например, вкусовых и слуховых рецепторах человека) раздражитель непосредственно воспринимается специализированными клетками эпителиального происхождения или видоизмененными нервными клетками (чувствительные элементы сетчатки), которые не генерируют нервных импульсов, а действуют на иннервирующие их нервные окончания, изменяя секрецию медиатора. В других случаях единственным клеточным элементом рецепторного комплекса является само нервное окончание, часто связанное со специальными структурами межклеточного вещества (например, тельце Пачини).
В технических науках для обозначения воспринимающего элемента чаще используется термин сенсор или датчик (от англ. sensor или чувствительный), предназначенное для преобразования спроецированного на него оптического изображения в электрический сигнал и его сканирования.
Клеточные рецепторы
В многоклеточном организме передача информации между клетками, происходящая с участием гормонов, нейротрансмиттеров (медиаторов), нейропептидов и других биологически активных веществ, включает этап взаимодействия молекул этих веществ (их называют также лигандами) с соответствующими надмолекулярными структурами, или клеточными рецепторами. Они могут располагаться как внутри клетки (напр., клеточные Р. к стероидным гормонам, легко проникающим внутрь клетки благодаря их растворимости в липидах клеточной мембраны), так и на поверхности клеточной мембраны (клеточные Р. к белкам, пептидам, нейротрансмиттерам). Как внутриклеточные, так и мембранные клеточные Р. содержат центр связывания, обеспечивающий специфическое связывание лиганда с клеточными Р. После связывания, напр., молекулы стероидного гормона с цитоплазматическим Р. и образования комплекса гормон — клеточный Р. этот комплекс проникает внутрь клеточного ядра, где связывается с соответствующим акцептором, вслед за чем молекула гормона отделяется от комплекса и выходит в цитоплазму, при этом одновременно активируется генетический аппарат клетки (см.). Конечным итогом этой активации является резкое усиление синтеза ряда специфических и неспецифических белков клетки, что представляет собой ответ клетки-мишени на действие гормона.
Процессы, происходящие при связывании молекулы лиганда с клеточным Р., локализованным на клеточной мембране (см. Мембраны биологические), состоят из ряда этапов, протекающих с большой скоростью. Происходящее при этом изменение свойств фосфолипидного матрикса, окружающего клеточный Р., обеспечивает передачу сигнала от центра связывания лиганда (через ряд промежуточных звеньев) на аденилат-циклазный центр и его активацию. Циклическая АМФ (см. Аденозинфосфорные кислоты) является своеобразным вторым внутриклеточным переносчиком информации, определяющим ответ клетки на действие лиганда. Т. о., происходит активация соответствующих протеинкиназ, изменение проницаемости клеточной мембраны для ряда ионов, усиление экспрессии генетической информации. Важным открытием явилось обнаружение в ц. н. с. клеточных Р. к ряду нейрональных пептидов, напр, к группам пептидов, названных эндорфинами и энкефалинами, а также клеточных Р. ко многим психотропным лекарственным препаратам (имипрамину, галоперидолу, диазепаму и др.). Взаимодействие указанных выше лигандов с клеточными Р. изменяет способность нервных клеток отвечать на действие нейро-трансмиттеров, т. е. оказывает на их активность модулирующее действие. Напр., связывание препаратов бензодиазепинового ряда с бензодиазе-пиновыми клеточными Р. усиливает ответ ГАМК-ергических нейронов на действие гамма-аминомасляной к-ты (ГАМК), влияя в то же время на связывание ГАМК соответствующими клеточными Р. В связи с обнаружением в ц. н. с. эндогенных лигандов, конкурирующих с морфином за места связывания и обладающих морфиноподобным действием, проводится поиск эндогенных соединений типа эндогенного диазепама, эндогенного галоперидола и др., что может иметь в случае их обнаружения большое значение для клин, практики.
Нарушения механизмов рецепции играют важную роль в развитии ряда заболеваний человека, напр, нек-рых видов сахарного диабета, гиперхоли-стеринемии и др. Наряду с рассмотренными выше видами Р. на поверхности В- и Т-лимфоцитов выявлены мембранные клеточные Р., играющие важную роль в работе иммунной системы, а также клеточные Р. к ряду вирусов.
Библиография: Глебов Р. И. и Крыжановский Г. Н. Функциональная биохимия синапсов, М., 1978; Гранит Р. Электрофизиологическое исследование рецепции, пер. с англ., М., 1957; Розен В. Б. и Смирнов А. Н. Рецепторы и стероидные гормоны, М., 1981, библиогр.; Тамар Г. Основы сенсорной физиологии, пер. с англ., М., 1976; Физиология сенсорных систем, под ред. А. С. Батуева, с. 34, Л., 1976; Cell membrane receptors for drugs and hormones, a multidisciplinary approach, ed. by R. W. Straub a. L. Bolis, N. Y., 1978; Cell! membrane receptors for viruses, antigens, and antibodies, polypeptide hormones, and small molecules, ed. by R. F. Beers a. E. G. Bassett, N. Y., 1976; The receptors, a comprehensive treatise, ed. by R. D. O’Brien, v. 1, N. Y.— L., 1979.
Классификация рецепторов
Сенсорные системы
Высокоспециализированные рецепторные образования нервной системы, воспринимающие раздражения из внешней среды, от органов тела относятся к органам рецепции. Рецепторы преобразуют энергию раздражений в специфическую форму нервного возбуждения.
Совокупность нейронов, участвующих в восприятии раздражения, проведении возбуждения до коры головного мозга, её сенсорные клетки составляют единую систему – «анализатор» (И.П.
Павлов), иначе «сенсорную систему».
Сенсорная системавключает три отдела:
1) рецепторный – периферический, воспринимающий определённую форму физической и химической энергии и транспортирующий её в нервный процесс;
2) проводниковый – нервное возбуждение достигает промежуточных центров мозгового ствола, где устанавливается связь с различными эффекторными путями;
3) центральный – корковый (участки коры больших полушарий мозга, где нервное возбуждение качественно изменяется, воспринимается как ощущение).
Основные функции сенсорной системы
1) восприятие и обработка информации (физической и химической энергии);
2) трансформация энергии в нервные импульсы;
3) передача нервных импульсов в мозг через цепи нейронов;
4) осуществление обратных связей.
Классификация рецепторов по характеру взаимодействия раздражителей
а) Экстерорецепторы (внешние рецепторы) – воспринимают раздражение при действии предметов и явлений внешнего мира. К ним относятся: слуховые, зрительные, обонятельные, вкусовые, тактильные раздражители.
б) Интерорецепторы (внутренние рецепторы) – воспринимают раздражения от внутренних органов (висцерорецепторы), от положения тела и движения тела и отдельных его частей в пространстве (вестибулорецепторы и проприорецепторы).
В зависимости от физической природы раздражителей.
а) Фоторецепторы – рецепторы зрительной сенсорной системы.
б) Механорецепторы – рецепторы скелетно-мышечной системы, слуховой, вестибулярной, тактильной систем, барорецепторы сердечно-сосудистой системы.
в) Терморецепторы – рецепторы кожи и внутренних органов.
г) Хеморецепторы – рецепторы обонятельной и вкусовой сенсорных систем, рецепторы сосудистые и тканевые.
д) Болевые – рецепторы болевой сенсорной системы.
В зависимости от контакта с раздражителем.
а) Дистантные рецепторы – воспринимающие раздражение на удалённом расстоянии от организма (зрительные, слуховые, обонятельные).
б) Контактные рецепторы – когда раздражитель вступает с ними в непосредственный контакт (вкусовые, тактильные).
По структурным особенностям и принципу преобразования энергии раздражения.
а) Первичночувствующие – тела чувствительных биполярных нейронов, находящихся на периферии; один отросток – дендрит – подходит к воспринимающей раздражение поверхности, другой (центральный) – аксон – передаёт возбуждение в центр. Энергия раздражителя трансформируется в нервный импульс в одной и той же клетке.
Возникает рецепторный потенциал, обуславливающий появление потенциала действия (ПД), достигающий по нервному волокну центров обоняния, тактильной и проприоцептивной чувствительности.
б) Вторичночувствующие – рецепторы, у которых между окончаниями сенсорного нейрона и точкой приложения раздражителя имеется рецептирующая (специализированная) клетка ненервного происхождения.
Возникающее в ней возбуждение по существу является рецепторным потенциалом. Выделяется медиатор, который передаётся через синапс на сенсорный нейрон.
Виды некоторых рецепторов и их данные[править | править код]
Cм. в таблице:
| Типы рецепторов и их данные | ||
|---|---|---|
| Вид раздражителя | Тип рецептора | Комментарии |
| Электрическое поле | ||
| Хеморецептор | ||
| Относятся к осморецепторам или механорецепторам. Располагаются на антеннах и ротовых органах многих насекомых | ||
| Механорецептор | У человека имеются в коже (экстероцепторы кожи) и внутренних органах (барорецепторы, проприоцепторы) | |
| Давление. | Относятся к механорецепторам | |
| (или механорецепторы вестибулярного аппарата), реагируют на ускорения и вибрации при наклоне тела или головы. | ||
| В основном интерорецепторы; у человека имеются в гипоталамусе, а также, вероятно, в почках, стенках желудочно-кишечного тракта, возможно, в печени. Существуют данные о широком распространении осморецепторов во всех тканях организма | ||
| Свет, Цвет | Фоторецептор,Экстерорецептор | Участвуют в восприятии света и цвета |
| Температура | Реагируют на изменение температуры. У человека они имеются в коже и в гипоталамусе. | |
| Болевые рецепторы — свободные нервные окончания немиелинизированных волокон типа C или слабо миелинизированных волокон типа Aδ. | ||
| Магнитное поле | Точное расположение и строение неизвестны, наличие у многих групп животных доказано поведенческими экспериментами. |
Рецепторы человека[править | править код]
Рецепторы кожи
- Свободные нервные окончания — нервные окончания, состоящие только из конечных ветвлений осевого цилиндра. Располагаются в эпителии. Выступают в качестве терморецепторов, механорецепторов и ноцицепторов (то есть отвечают за восприятие изменения температуры, механических воздействий и болевые ощущения).
- Несвободные нервные окончания:
- Тельца Пачини — инкапсулированные рецепторы давления в округлой многослойной капсуле. Располагаются в подкожно-жировой клетчатке. Являются быстроадаптирующимися (реагируют только в момент начала воздействия), то есть регистрируют силу давления. Обладают большими рецептивными полями, а потому обладают грубой чувствительностью.
- Тельца Мейснера — инкапсулированные рецепторы давления, расположенные в дерме. Представляют собой слоистую структуру с нервным окончанием, проходящим между слоями. Являются быстроадаптирующимися. Обладают малыми рецептивными полями, а потому обладают тонкой чувствительностью.
- Тельца Меркеля — некапсулированные рецепторы давления. Располагаются у птиц — в дерме, у прочих позвоночных — в глубоких слоях эпидермиса. Являются медленноадаптирующимися (реагируют на всей продолжительности воздействия), то есть регистрируют продолжительность давления. Обладают малыми рецептивными полями.
- Тельца Руффини — инкапсулированные рецепторы растяжения. Являются медленноадаптирующимися, обладают большими рецептивными полями. Реагируют также на тепло.
- Колбы Краузе — инкапсулированные рецепторы, расположенные в надсосочковом слое дермы. Раньше считалось, что у Колб Краузе есть специфическая чувствительность, но их роль в качестве холодовых рецепторов не подтвердилась. .
- Рецепторы волосяных фолликулов — механорецепторы, расположенные в волосяных фолликулах и реагирующие на отклонение волоса от исходного положения.
Рецепторы мышц и сухожилий (проприоцепторы)
- Мышечные веретена — рецепторы растяжения мышц, бывают двух типов:
- с ядерной сумкой
- с ядерной цепочкой
- Сухожильный орган Гольджи — рецепторы сокращения мышц. При сокращении мышцы сухожилие растягивается и его волокна пережимают рецепторное окончание, активируя его.
Рецепторы связок
В основном представляют собой свободные нервные окончания (Типы 1, 3 и 4), меньшая группа — инкапсулированные (Тип 2). Тип 1 аналогичен окончаниям Руффини, Тип 2 — тельцам Паччини.
Рецепторы сетчатки глаза
Сетчатка содержит палочковые и колбочковые фоточувствительные клетки, в которых имеются светочувствительные пигменты. Палочки чувствительны к очень слабому свету, это длинные и тонкие клетки, сориентированные по оси прохождения света. Все палочки содержат один и тот же светочувствительный пигмент. Колбочки требуют намного более яркого освещения, это короткие конусообразные клетки, у человека колбочки делятся на три вида, каждый из которых содержит свой светочувствительный пигмент — это и есть основа цветового зрения.
Под воздействием света в рецепторах происходит выцветание — молекула зрительного пигмента поглощает фотон и превращается в другое соединение, хуже поглощающее свет на этой длине волны. Практически у всех животных (от насекомых до человека) этот пигмент состоит из белка, к которому присоединена небольшая молекула, близкая по структуре к витамину A. Эта молекула и представляет собой химически трансформируемую светом часть. Белковая часть выцветшей молекулы зрительного пигмента активирует молекулы трансдуцина, каждая из которых деактивирует сотни молекул циклического гуанозинмонофосфата, участвующих в открытии пор мембраны для ионов натрия, в результате чего поток ионов прекращается — мембрана гиперполяризуется.
Чувствительность палочек такова, что адаптировавшийся к полной темноте человек способен увидеть вспышку света такую слабую, что каждый рецептор получит не больше одного фотона. При этом палочки не способны реагировать на изменения освещённости, когда свет настолько ярок, что все натриевые каналы уже закрыты.
Виды некоторых рецепторов и их данные
Cм. в таблице:
| Типы рецепторов и их данные | ||
|---|---|---|
| Вид раздражителя | Тип рецептора | Комментарии |
| Электрическое поле | Ампула Лоренцини и другие типы | Имеются у рыб, круглоротых, амфибий, а также у утконоса и ехидны |
| Химическое вещество | Хеморецептор | Химический сигнал преобразуется в потенциал действия. Хеморецепторы (вкуса, запаха, феромонов) могут быть очень различны по своей природе, и потому по-разному реагировать на одно и то же вещество — в зависимости от точки его приложения (нос, рот, кожа и т. п.). |
|
Влажность
ПММА |
Гигрорецептор | Относятся к осморецепторам или механорецепторам. Располагаются на антеннах и ротовых органах многих насекомых |
| Механическое воздействие. | Механорецептор | У человека имеются в коже (экстероцепторы кожи) и внутренних органах (барорецепторы, проприоцепторы) |
| Давление. | Барорецептор | Относятся к механорецепторам |
| Положение тела | Проприоцептор | нервно-мышечные веретена, сухожильные органы Гольджи и др. |
| Положение тела | Вестибулорецептор | (или механорецепторы вестибулярного аппарата), реагируют на ускорения и вибрации при наклоне тела или головы. |
| Осмотическое давление | Осморецептор | В основном интерорецепторы; у человека имеются в гипоталамусе, а также, вероятно, в почках, стенках желудочно-кишечного тракта, возможно, в печени. Существуют данные о широком распространении осморецепторов во всех тканях организма |
| Свет, Цвет | Фоторецептор,Экстерорецептор | Участвуют в восприятии света и цвета |
| Температура | Терморецептор | Реагируют на изменение температуры. У человека они имеются в коже и в гипоталамусе. |
| Повреждение тканей | Ноцицептор | Болевые рецепторы — свободные нервные окончания немиелинизированных волокон типа C или слабо миелинизированных волокон типа Aδ. |
| Магнитное поле | Магнитные рецепторы | Точное расположение и строение неизвестны, наличие у многих групп животных доказано поведенческими экспериментами. |
Лиганд (биохимия)
Основная статья: Лиганд (биохимия)
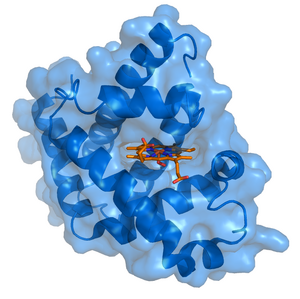
Миоглобин (голубой) со связанным лигандом — гемом (оранжевый). Данные из Protein Data Bank
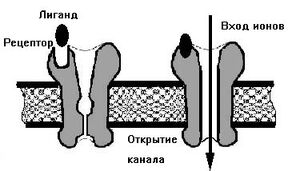
Рис.4. Работа лиганда.
Лиганд (биохимия) (Биохимия и Фармакология) — это химическое соединение (часто малая молекула), которое образует комплекс (в химии) с той или иной биомолекулой (чаще белком, например клеточным рецептором, но иногда, например, с ДНК) и производит, в результате такого связывания, те или иные биохимические, физиологические или фармакологические эффекты. В случае связывания лиганда с белком, лиганд обычно является малой сигнальной молекулой, связывающейся со специфическим участком связывания на белке-мишени (например, на рецепторе), (смотри работу лиганда рис.4.). В случае связывания лиганда с ДНК, лиганд обычно также является малой молекулой или ионом, или белком который связывается с двойной спиралью ДНК.
Лаборатория клеточной биологии рецепторов РАН

Уникальная научная установка «Система зондово-оптической 3D — трёхмерной корреляционной микроскопии»
Лаборатория, оснащённая современным оборудованием, занимается изучением механизмов функционирования клеточных рецепторов. В настоящее время проводится работа по двум основным направлениям:
Первое связано с исследованием адгезионного нейронального G-белоксопряженного рецептора CIRL. Рецепторы CIRL представляют собой природные гибриды двух классов белков – сигнальных рецепторов и молекул клеточной адгезии. Считается, что эти химерные рецепторы могут быть вовлечены в межклеточные взаимодействия и передачу сигналов, опосредованных G-белками. Однако до сих пор для рецепторов данного семейства не были найдены природные агонисты.
Второе направление связано с исследованием представителя семейства инсулинового рецептора – IRR (insulin receptor-related receptor). К этому семейству принадлежат также инсулиновый рецептор (IR) и рецептор инсулино-подобного фактора роста (IGF-IR). Лигандами рецепторов IR и IGF-IR являются эндогенные пептиды, тогда как для IRR до недавнего времени не удавалось обнаружить лиганд, несмотря на значительные усилия, предпринятые в этом направлении.
Системы вторых посредников
цАМФ. В интеграции внешних сигналов участвуют также системы вторых посредников. Хотя рецепторов и белковых сигнальных молекул известно гораздо больше, чем вторых посредников, последние задействованы во множестве путей внутриклеточной передачи сигнала. К наиболее изученным вторым посредникам относятся цАМФ, цГМФ, Са2+, ИФ3, ДАГ, N0. Эта группа разнородных соединений постоянно пополняется. Вторые посредники взаимодействуют напрямую (изменяя метаболизм друг друга) или косвенно (воздействуя на одни и те же внутриклеточные мишени). Благодаря таким на первый взгляд сложным регуляторным механизмам отдельные внешние сигналы или их сочетание приводят в действие интегрированные системы вторых посредников, вызывая согласованную клеточную реакцию. Функцию вторых посредников, а также регуляцию их образования (или высвобождения), расщепления и выведения из клетки удобно рассматривать на примере цАМФ. Этот второй посредник синтезируется под действием аденилатциклазы при активации многих рецепторов, сопряженных с G-белками. Gj-белок активирует аденилатциклазу, Gj-белок — ингибирует.
Существует не менее десяти тканеспецифичных изоформ аденилатциклазы, различающихся по механизмам регуляции активности. Некоторые изоформы активируются или ингибируются комплексом из β- и у-субъединиц G-белка. Поскольку эти субъединицы общие у всех G-белков, активность аденилатциклазы могут регулировать не только Gr и Gj-белки, но и другие типы G-белков. Другие изоформы стимулируются Са2+ или Са2+-кальмодулиновым комплексом. Разница в механизмах регуляции активности аденилатциклазы (они могут включать фос-форилирование и другие регуляторные влияния) обеспечивает широкий спектр регуляторных влияний на разные клетки-мишени. цАМФ гидролизуется под действием фосфодиэстераз и выводится из клетки с помощью нескольких мембранных транспортных белков. Фосфодиэстеразы относятся к белковым сигнальным молекулам; их активность регулируется на уровне транскрипции, а также при взаимодействии со вторыми посредниками (цАМФ, цГМФ, Са2+) и другими сигнальными белками.
Как правило, цАМФ активирует протеин киназы А (цАМФ-зависимые протеинкиназы) — небольшую группу родственных белков. Эти протеинкиназы, в свою очередь, фосфорилируют не только конечные внутриклеточные мишени (ферменты, транспортные белки), но и другие протеинкиназы и прочие регуляторные белки. К последним относятся, например, факторы транскрипции. Они отвечают за опосредованную цАМФ регуляцию транскрипции генов, обеспечивая отсроченную клеточную реакцию на сигнал. Помимо активации протеинкиназ цАМФ воздействует непосредственно на мембранные катионные каналы, играющие важную роль, в частности, в функционировании обонятельных нейронов. Таким образом, сигнал от цАМФ вызывает цепь биохимических изменений в клетке-мишени.
Кальций. Еще один хорошо изученный второй посредник — внутриклеточный Са2+. Ионы Са2+ поступают в цитоплазму разными путями: по мембранным каналам (зависимым от G-белков, потенциалзависимым, регулируемым К+ либо самим Са +), а также по каналам, расположенным в особых участках эндоплазматического ретикулума и открывающимся под действием ИФ3, а в скелетных мышцах — в результате деполяризации мембраны. Удаление кальция из цитоплазмы происходит двумя путями: он поглощается эндоплазматическим ретикулумом или выводится из клетки. Са2+ передает сигналы гораздо большему числу белков, чем цАМФ — ферментам, участвующим в клеточном метаболизме, протеинкиназам, кальцийсвязывающим белкам. Последние взаимодействуют с другими конечными и промежуточными эффекторами.
Антиноцицептивные системы и рецепторы
В основу классификации рецепторов положены следующие принципы:
1. Среда, в которой рецепторы воспринимают информацию (экстеро-, интеро-, проприо- и другие рецепторы).
Экстерорецепторы воспринимают раздражения внешних агентов (рецепторы органов слуха, зрения, обоняния, вкуса, осязания).
Интерорецепторы сигнализируют о раздражителях внутренней среды (рецепторы внутренних органов).
Проприоцепторы (рецепторы опорно-двигательного аппарата).
Природа адекватного раздражителя (механо-, термо-, фото- и другие рецепторы).
Механорецепторы приспособлены к восприятию механической энергии раздражающего стимула.
Терморецепторы воспринимают температурные раздражения.
Хеморецепторы чувствительны к действию химических агентов.
Фоторецепторы воспринимают световую энергию
3. Характер ощущения после контакта с рецепторами (тепловые, холодовые, болевые и др.).
Способность воспринимать раздражитель, находящийся на расстоянии от рецептора — дистантный (обонятельный, зрительный) или при непосредственном контакте с ним — контактный (вкусовой, тактильный).
По количеству воспринимаемых модальностей (раздражителей) рецепторы могут быть мономодальными (например, световой) и полимодальными (механический и температурный).
Морфологические особенности и механизмы возникновения возбуждения. Различают первичночувствующие (обонятельные, тактильные) и вторичночувствующие рецепторы (зрения, слуха, вкуса).
Первичночувствующие рецепторы — это биполярные сенсорные нервные клетки, снабженные ресничками, которые наподобие антенн ведут «поиск» адекватного раздражителя.
Контакт с раздражителем приводит к возникновению рецепторного потенциала, который электротонически распространяется к аксону сенсорного нейрона, где формируется ПД, распространяющийся по нервному волокну.
К вторичночувствующим рецепторам относятся те рецепторы, у которых между сенсорными нейронами и раздражителем существует еще дополнительная рецептирующая клетка нервного происхождения, например, палочки и колбочки, волосковые клетки слухового анализатора.
После контакта с раздражителем в рецептирующей клетке возникает рецепторный потенциал, который с помощью синаптической передачи между рецептирующей клеткой и нервным волокном сенсорного нейрона способствует возникновению в нем генераторного потенциала. Последний на аксоне нейрона преобразуется в ПД, который электротонически распространяется по нервному волокну.
При отношении биологической системы к видам раздражителей (света, звука, механической силы, химических факторов) они разделяются на две группы: адекватные и неадекватные.
Адекватные – это те, к которым рецепторный аппарат клетки на протяжении эволюции приспособился путем повышения возбудимости (порог возбуждения у них низкий).
Неадекватные – раздражители, которые не отвечают биологическим особенностям ткани.
Они вызывают возбуждение только тогда когда появляется повреждение в биологической системе.
Механизмы возбуждения рецепторов. Первичные (первичночувствующие) и вторичные (вторичночувствующие) рецепторы.
Визуализация эффектов адреналина
После успешного выделения гена, Брайан Кобилка перебрался на Медицинский факультет Стэнфордского университета в Калифорнии. Там он намеревался определить структуру рецептора, что, по мнению большинства в научном сообществе на тот момент, было совершенно недостижимой целью. И это стало для Кобилки занятием надолго.
Визуализация структуры белка — процесс, включающий в себя много сложных этапов. Белки слишком малы, чтобы их можно было разглядеть в обычный микроскоп, поэтому ученые используют метод, называемый рентгеновской кристаллографией. Они начинают с выращивания кристаллов, в которых молекулы белка плотно упакованы в виде симметричной решетки подобно тому, как молекулы воды упакованы в кристалле льда или атомы углерода в алмазе. Затем исследователи подвергают кристаллы действию рентгеновского излучения. Когда лучи ударяют по молекулам белка, они рассеиваются, и по характеру получающейся картины рассеяния ученые могут рассчитать, как выглядят белки на атомарном уровне. Первое изображение кристаллической структуры белка было получено в 1950-х годах, и с тех пор ученым удалось получить структуры тысяч белков. Однако большая часть из них относится к водорастворимым белкам, для которых вырастить кристалл относительно легко. Гораздо меньшему числу исследователей удалось получить изображение белков, расположенных в липидных мембранах клеток . Более того, рецепторы из семейства GPCR по своей природе очень подвижные (они передают сигнал посредством внутренних перестроек), но внутри кристалла они должны стать почти полностью статичными. Таким образом, уже сама по себе их кристаллизация является непростой задачей.
О структурных исследованиях белковых молекул мы писали в статье «Ловля бабочек, или чем структурная геномика поможет биологии» . О роли липидных молекул в формировании мембраны и в самой жизни читайте в статье «Липидный фундамент жизни» . — Ред.
Кобилке потребовалось более двух десятилетий, чтобы найти решение этих задач. Но, благодаря решительности, творческому подходу и молекулярно-биологическим приемам, Кобилка и его группа достигла, наконец, заветной цели в 2011 году: они получили изображение рецептора в тот самый момент, когда он передает сигнал от гормона снаружи клетки G-белку внутри нее (рис. 3) . Эта структура, опубликованная в журнале Nature, открыла новые подробности о рецепторах GPCR. Например, стало понятно, как выглядит активированный рецептор, когда он открывает полость для связывания G-белка (рис. 4). Такая информация окажется очень полезной в будущем для разработки новых фармацевтических препаратов.
О первом успехе лаборатории Кобилки в структурной биологии GPCR-рецепторов «биомолекула» писала в статье «Новый рубеж: получена пространственная структура β2-адренорецептора» . Первая работа, в которой удалось «увидеть» активированный рецептор, описана в заметке «Рецепторы в активной форме» . — Ред.
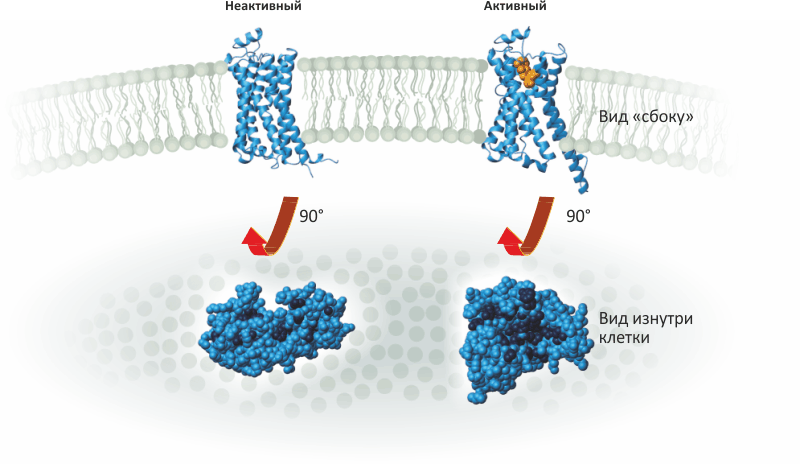
Рисунок 4. β-Адренергический рецептор при активации изменяет форму. Когда гормон (показан оранжевым) присоединяется снаружи рецептора, его (рецептора) внутренняя часть раскрывается подобно букету цветов. На картинке внизу рецептор повернут так, что видна его внутриклеточная сторона. Водорастворимая часть рецептора показана голубым; гидрофобные фрагменты, выступающие на поверхность лишь при активации, показаны темно-синим. Когда гормон связывается (справа), открывается гидрофобная полость, с которой и связывается α-субъединица G-белка.
Выманивание рецепторов из потаенных мест
В 1970 году Лефковиц публикует статьи в двух престижных журналах Proceedings of the National Academy of Sciences (PNAS) и Science, в которых описывает открытие активного рецептора. Это достижение дает ему почувствовать волнение исследовательской работы, и в итоге он оказывается в Дюкском университете в Северной Каролине. Не то чтобы он туда особенно стремился, но там ему сделали предложение, от которого он просто не мог отказаться. В новой лаборатории Лефковиц собирает свою собственную команду исследователей. И хотя уже было ясно, что он никогда не станет кардиологом, он все равно хочет заниматься болезнями сердца.
Итак, он концентрируется на рецепторах адреналина и норадреналина, так называемых адренергических рецепторах. Используя радиоактивно меченные вещества, включая бета-блокаторы, его группа изучает работу этих рецепторов, и после прецизионной настройки оборудования удается выделить ряд рецепторов из биологической ткани. Между тем, возросло понимание того, что происходит внутри клетки. Исследователи обнаружили то, что они назвали G-белками (Нобелевская премия по физиологии и медицине за 1994 год) и что активируется по сигналу со стороны рецепторов. В свою очередь, G-белок запускает цепь реакций, которые изменяют метаболизм клетки. С начала 1980-х к ученым стало приходить понимание пути, по которому сигнал передается снаружи клетки вовнутрь (рис. 2).
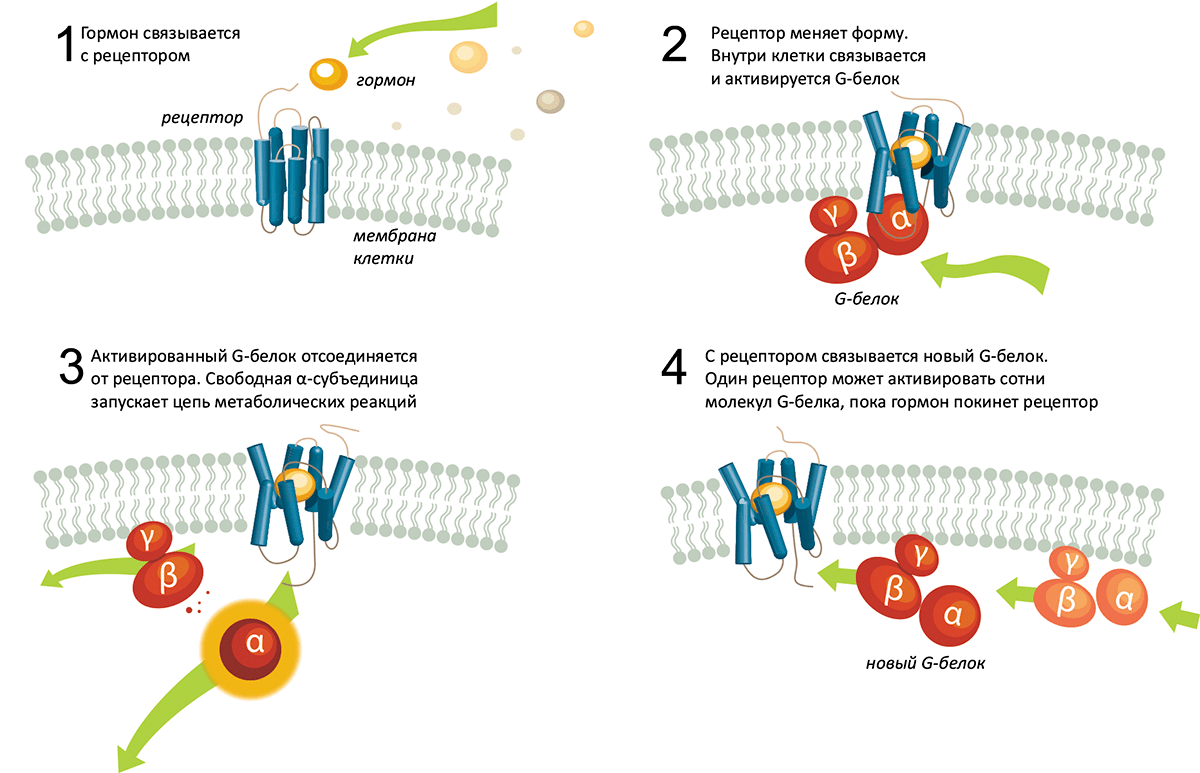
Рисунок 2. Когда гормон, «молекула с запахом» или «молекула со вкусом» связываются с рецептором на поверхности клетки, запускается каскад реакций внутри неё
Секретные PRR
Некоторые PRR не остаются связанными с клеткой, которая их производит. Рецепторы комплемента , коллектины , фиколины , пентраксины, такие как сывороточный амилоид и С-реактивный белок , липидные трансферазы , белки распознавания пептидогликана (PGRP) и LRR, XA21D, все являются секретируемыми белками. Одним из очень важных коллектинов является маннан-связывающий лектин (MBL), основной PRR врожденной иммунной системы, которая связывается с широким спектром бактерий, вирусов, грибов и простейших. MBL преимущественно распознает определенные группы сахаров на поверхности микроорганизмов, но также связывает фосфолипиды , нуклеиновые кислоты и негликозилированные белки. После связывания с лигандами олигомеры MBL и фиколина рекрутируют MASP1 и MASP2 и инициируют лектиновый путь активации комплемента, который в некоторой степени похож на классический путь комплемента .






