Башкирский государственный медицинский университет
Содержание:
- Классификация
- Приложения
- Презентация на тему: » Cинаптическая передача нервного импульса. Место контакта, передачи нервного импульса Передача нервного возбуждения может осуществляться электрически (через.» — Транскрипт:
- Структура [ править ]
- Гладкие мышцы
- Суммирование [ править ]
- Физиология мышечных волокон
- Связь с электрическими синапсами [ править ]
- Объемная передача [ править ]
- Нервно-мышечное соединение
- Синаптический уровень
- Механизм сокращения поперечно-полосатых мышц
- Внешние ссылки [ править ]
- Интеграция сигналов: увеличьте постсинаптический элемент
- Действие наркотиков [ править ]
- Особенности проведения возбуждения в нервно-мышечном синапсе
- Механизм синаптической передачи
- Cotransmission [ править ]
Классификация
На уровне сомы:
— Простой аксосоматический синапс,
— Инвагинированный аксосоматический синапс,
— Остистый аксосоматический синапс.
— простой аксодендритный синапс (соединение двух мембран),
— Шипастый аксодендритный синапс (синапс на зачатке дендритных шипов),
— гребенчатый синапс (на стороне дендритного зачатка),
— разветвленный колючий синапс,
— Аксо — дендритный синапс «попутно»,
— реципрокный аксодендритный синапс,
— полисинаптическое окончание,
— Межпальцевой колючий синапс.
На уровне конуса имплантации и аксона:
— Проксимальный аксо — аксонный синапс,
— Тормозной аксо — аксонный синапс,
— Дистальный аксо — аксонный синапс,
— Аксо — аксонный синапс «между прочим»,
Дендритные окончания сверхмобильны.
Приложения
О других проектах Викимедиа:
Библиография
Дейл Первес, Джордж Дж. Августин, Дэвид Фицпатрик, Уильям К. Холл, Энтони-Самуэль Ламантия, Джеймс О. Макнамара и С. Марк Уильямс, нейронауки , Брюссель, Университет Де Бока , колл. «Неврология и познание»,2005 г., 3 е изд. , 811 с. ( ISBN 978-2-8041-4797-6 , читать онлайн ) , гл. 5 («Синаптическая передача»), с. 93-127.
- Глиальная клетка
- Синаптическая обрезка
- Синаптическая пластичность
- Постсинаптический потенциал
- Рефлекс разгибания
- Сгибательный рефлекс
- Мозг
- Спинной мозг
- Тематическая классификация нейробиологии
- История электрофизиологии
- Электрофизиология
- Потенциал отдыха
- Биофизика ионных каналов
- Взаимный синапс
Внешние ссылки
-
Авторитетные записи :
- ( )
- Ресурсы, связанные со здоровьем :
- (la + en) TA98
|
Нейронауки |
||
|---|---|---|
| Основополагающие исследования |
|
|
| Клиническая неврология |
|
|
| Когнитивная нейробиология |
|
|
| Методы |
|
|
| Основные концепции |
|
|
| Исследователи |
|
|
| Междисциплинарные области |
|
|
| Список анатомических устройств и человеческих систем |
|
Физиология нервной системы |
|||||||||
|---|---|---|---|---|---|---|---|---|---|
| Центральная нервная система (ЦНС) | Физиологическая активация · Состояние сознания · Внутричерепная гипертензия · Асимметрия мозга · Сон · Память | ||||||||
| Периферическая нервная система (ПНС) | Рефлекс · Ощущение | ||||||||
| Оба |
|
- Портал нейробиологии
- Портал физиологии
- Портал клеточной и молекулярной биологии
Презентация на тему: » Cинаптическая передача нервного импульса. Место контакта, передачи нервного импульса Передача нервного возбуждения может осуществляться электрически (через.» — Транскрипт:
1
Cинаптическая передача нервного импульса

2
Место контакта, передачи нервного импульса Передача нервного возбуждения может осуществляться электрически (через электрический синапс) или с помощью химического посредника – медиатора (через химический синапс) Синапс – это место контакта между окончанием аксона и мембраной другой нервной или рабочей клетки.
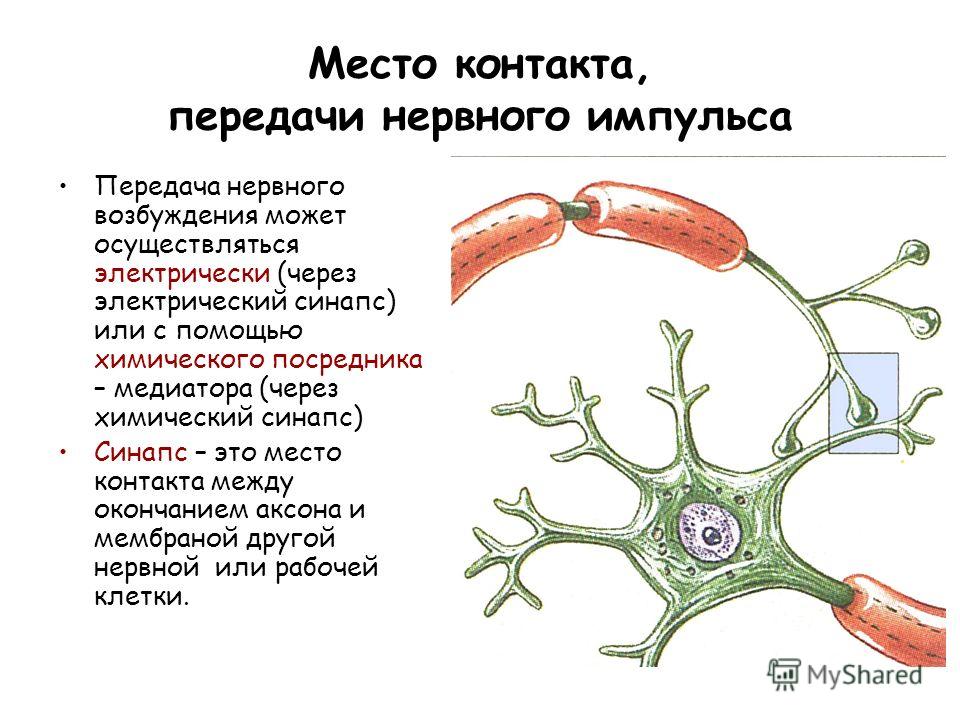
3
Плотность расположения синапсов на нервной клетке 1 –тело нервной клетки 2 – синапсы на дендритах 3 – аксон 4 — дендриты

4
Классификация синапсов По локализации: центральные (головной и спинной мозг) и перифенические. По конечному эффекту: тормозные и возбуждающие По механизму передачи сигнала: электрические, химические, смешанные По природе медиатора: холинергические (медиатор ацетилхолин), адренергические (медиатор норадреналин), ГАМК-ергические (медиатор гамма-аминомасляная кислота) и др.

5
Химический синапс — место передачи нервного импульса Химические синапсы наиболее распространены в ЦНС. В химический синапс входят: синаптический холмик с пресинаптической мембраной, синаптическая щель, постсинаптическая мембрана. Медиатор (посредник) находится в пузырьках нервного окончания.
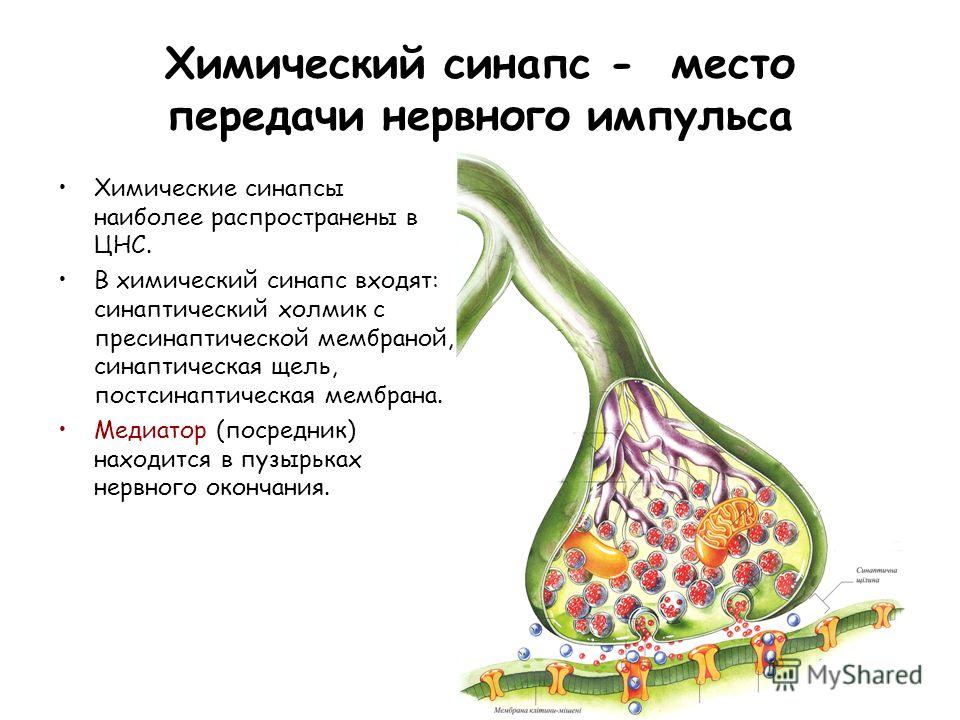
6
Выделение медиатора 1.ПД вызывает деполяризацию пресинаптической мембраны, что увеличивает вход в пресинаапс ионов кальция. 2.В ответ высвобождаются кванты медиатора. 3. Пройдя через синаптическую щель медиатор взаимодействует с рецептором на постсинаптической мембране. 4. Это сопровождается изменением ионной проводимости постсинаптической мембраны. 5. Если повышается проницаемость для ионов натрия – возникает деполяризация – возбуждающий синапс. 6. Если повышается проницаемость для калия – возникает гиперполяризация – тормозной синапс
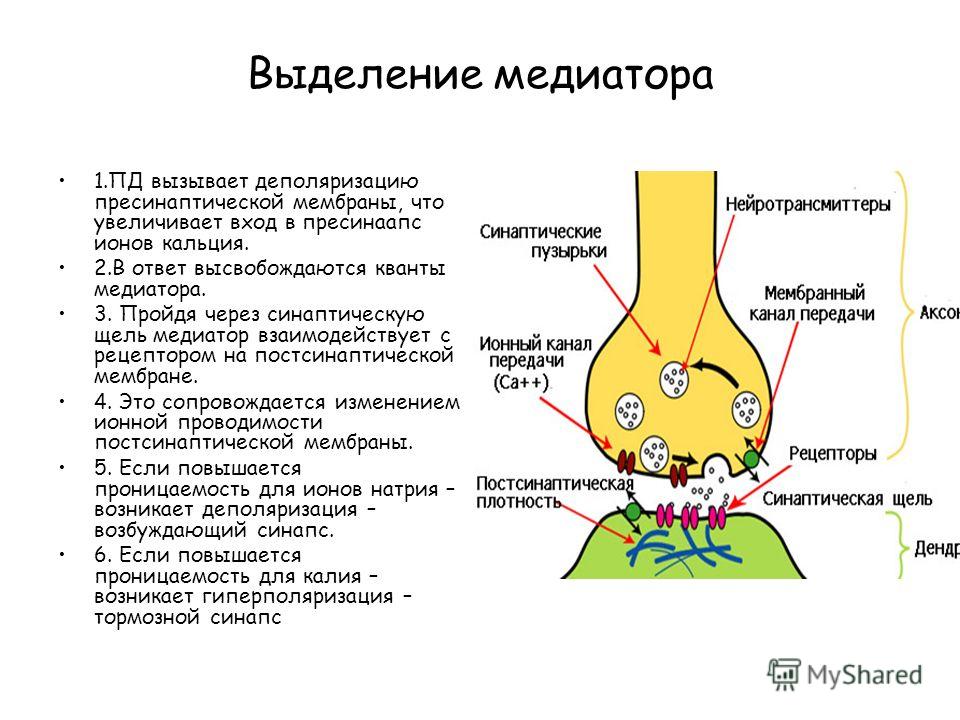
7
Деполяризация постсинаптической мембраны синапса Каждый квант медиатора вызывает развитие миниатюрного потенциала. ПД возникает тогда, когда одновременно или последовательно с короткими промежутками времени выделяется несколько квантов медиатора и достигается критический уровень деполяризации.
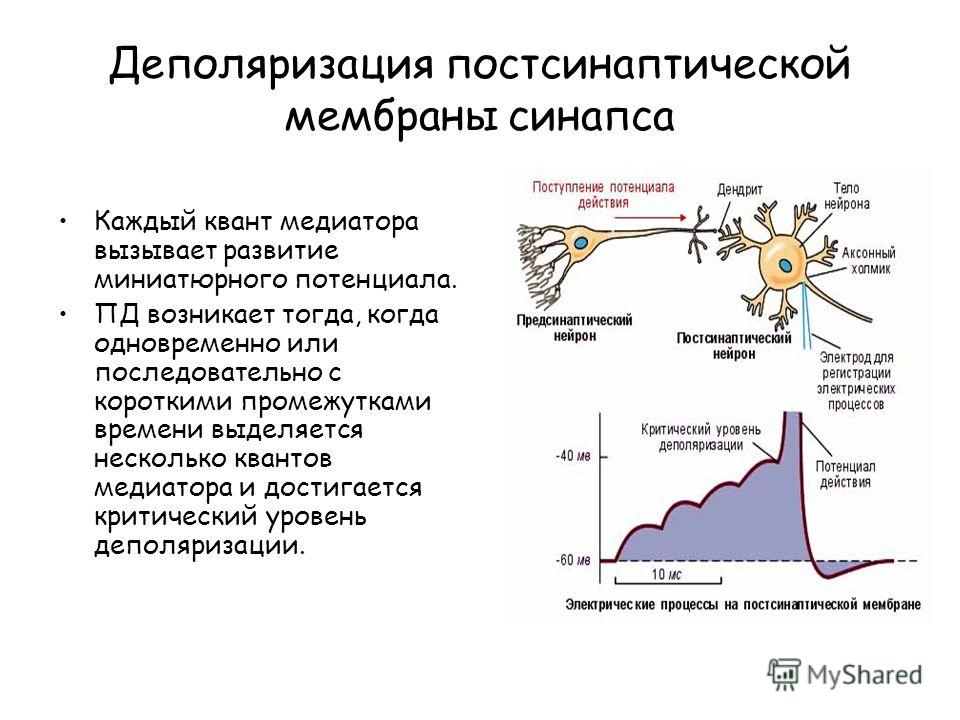
9
Свойства химических синапсов 1. Обеспечивают одностороннюю передачу возбуждения 2.Имеется синаптическая задержка (0,2 – 0,5 мс) 3. Обеспечивают как возбуждение, так и торможение 4. Сохраняют следы предшествующей активности (имеет значение для формирования памяти) 5. Чувствительны к изменению температуры

10
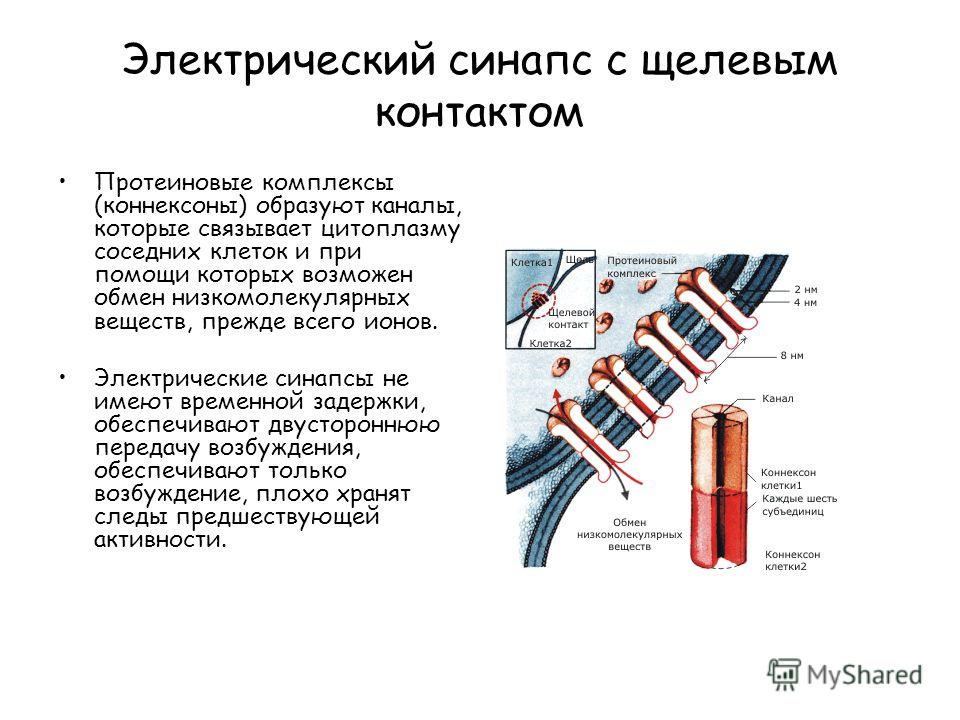
Электрический синапс с щелевым контактом Протеиновые комплексы (коннексоны) образуют каналы, которые связывает цитоплазму соседних клеток и при помощи которых возможен обмен низкомолекулярных веществ, прежде всего ионов. Электрические синапсы не имеют временной задержки, обеспечивают двустороннюю передачу возбуждения, обеспечивают только возбуждение, плохо хранят следы предшествующей активности.
11
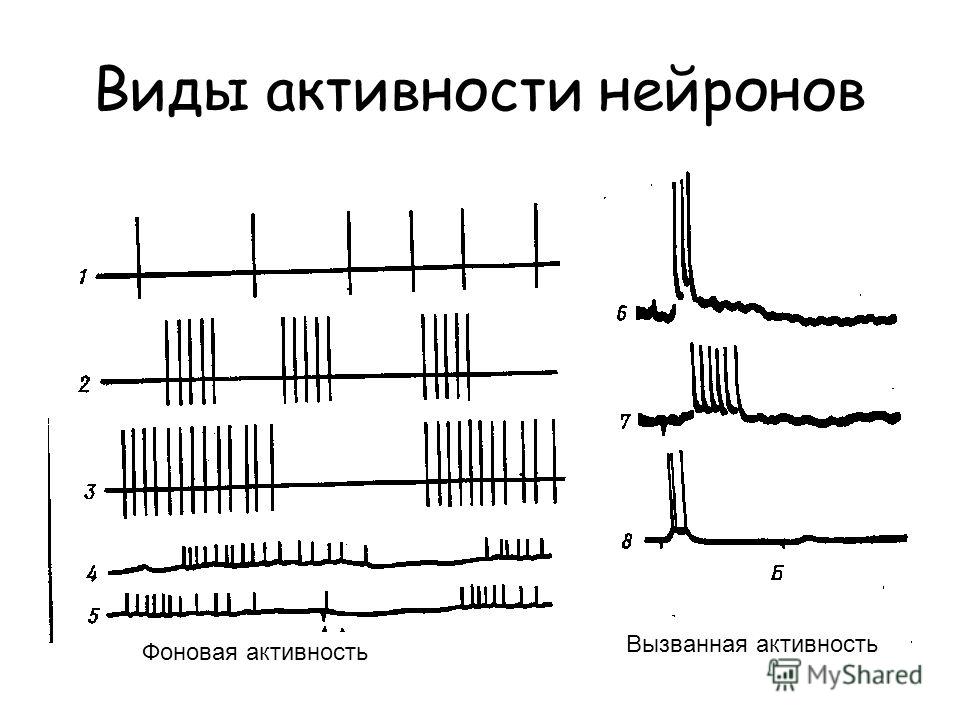
Виды активности нейронов Фоновая активность Вызванная активность
12
Химические посредники — медиаторы Основные медиаторы: Амины: :ацетилхолин (АХ), норадреналин (НА) или норэпинефрин, Дофамин, серотонин. Аминокислоты: глицин, глутамин, ГАМК – гамма- аминомасляная кислота и др. Пуриновые нуклеотиды – АТФ Нейропептиды: гипоталамические либерины и статины, опиоиды, субстанция Р и др.

13
Ацетилхолин-медиатор Ацетилхолин образуется из холина и ацетилхоэнзима А в цитоплазме окончаний холинергических нервов. Есть два вида холинорецепторов: М — мускариночувствительные (чувствительны к яду гриба мухомора) и Н –никотинчувствительные (к никотину). АХ после выделения разрушается почти мгновенно ферментом холинэстеразой. ЯДЫ: ботулин – блокирует синтез и выход АХ Кураре – блокирует холинорецепторы скелетных мышц ФОС (фосфорорганические соединенияч) блокируют активность холинэстеразы Атропин – блокирует М-холинорецепторы

14
Норадреналин-медиатор Образуется из дофамина. Выделяют несколько видов рецепторов: альфа (α 1, α 2 ) и несколько видов бета. После выделения разрушается ферментом моноаминооксидазой. Яды: резерпин – нарушает синтез НА Пирогаллол – угнетает активность МАО Эфедрин – усиливает выделение НА Обзидан – бета-адреноблокатор Кокаин – угнетение обратного захвапта НА

Структура [ править ]
| Структура типичного химического синапса |
|---|
|
Постсинаптическая плотность Вольт — закрытого типа Ca ++ канала Синаптический пузырек Транспортер нейротрансмиттера Рецептор Нейротрансмиттер Аксон терминал Синаптическая щель Дендрит |
| Различие между до и после синапса |
|---|
|
↓ К постсинаптическому нейрону ↓ Из пресинаптического нейрона Транспортер нейротрансмиттера Рецептор нейротрансмиттера Передача нейротрансмиттера Синаптическая щель { |
| «Соединение, связывающее нейрон с нейроном, — это синапс. Сигнал течет в одном направлении, от пресинаптического нейрона к постсинаптическому нейрону через синапс, который действует как переменный аттенюатор». Вкратце, направление потока сигнала определяет префикс для задействованных синапсов. |
Синапсы — это функциональные связи между нейронами или между нейронами и другими типами клеток. Типичный нейрон дает начало нескольким тысячам синапсов, хотя есть некоторые типы, которые образуют гораздо меньше. Большинство синапсов соединяют аксоны с дендритами , , но есть и другие типы соединений, включая аксон-к-телу клетки, аксон-аксон, и от дендрита к дендриту . Синапсы обычно слишком малы, чтобы их можно было распознать с помощью светового микроскопа.за исключением точек, где кажется, что мембраны двух клеток соприкасаются, но их клеточные элементы можно четко визуализировать с помощью электронного микроскопа .
Химические синапсы направленно передают информацию от пресинаптической клетки к постсинаптической клетке и поэтому асимметричны по структуре и функциям. Пресинаптического аксона терминал или синаптическойбутон — это специализированная область в аксоне пресинаптической клетки, которая содержит нейротрансмиттеры, заключенные в небольшие мембраносвязанные сферы, называемые синаптическими пузырьками (а также ряд других поддерживающих структур и органелл, таких как митохондрии и эндоплазматический ретикулум ). Синаптические везикулы состыкованы с пресинаптической плазматической мембраной в областях, называемых активными зонами .
Сразу напротив находится область постсинаптической клетки, содержащая рецепторы нейротрансмиттеров ; для синапсов между двумя нейронами постсинаптическая область может находиться на дендритах или теле клетки. Сразу за постсинаптической мембраной находится сложный комплекс взаимосвязанных белков, называемый постсинаптической плотностью (PSD).
Белки в PSD участвуют в закреплении и транспортировке рецепторов нейротрансмиттеров, а также в модуляции активности этих рецепторов. Рецепторы и PSD часто находятся в специализированных выступах основного дендритного вала, называемых дендритными шипами .
Синапсы можно описать как симметричные или асимметричные. При исследовании под электронным микроскопом асимметричные синапсы характеризуются округлыми пузырьками в пресинаптической клетке и заметной постсинаптической плотностью. Асимметричные синапсы обычно возбуждающие. Симметричные синапсы, напротив, имеют уплощенные или удлиненные пузырьки и не содержат заметной постсинаптической плотности. Симметричные синапсы обычно тормозят.
Синаптическая щель -также называется синаптической щели — зазор между пре- и постсинаптических клеток , что составляет около 20 нм (0,02 мкм) в ширину. Небольшой объем щели позволяет быстро повышать и понижать концентрацию нейромедиаторов.
Autapse представляет собой химические (или электрические) синапсы образуются , когда аксон одного нейрона синапсов с его собственными дендритами.
Гладкие мышцы
Гладкие мышцы — это мышцы, формирующие слой стенок полых внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток без поперечной исчерченности за счет хаотичного расположения миофибрилл.
Особенности гладких мышц:
- Иннервируются волокнами вегетативной нервной системы (ВНС);
- Обладают низкой возбудимостью:
- Обладают низкой величиной МП (мембранного потенциала) — -50 — -60 мВ из-за более высокой проницаемости для ионов Na+
- ПД (потенциал действия) отличается меньшей амплитудой и большей длительностью. Он формируется в основном за счет ионов Ca2+
- Медленная проводимость:
Клетки в гладких мышцах функционально связаны между собой посредством щелевидных контактов — нексусов, которые имеют низкое электрическое сопротивление. За счет этих контактов ПД распространяется с одного мышечного волокна на другое, охватывая большие мышечные пласты, и в реакцию вовлекается вся мышца.
Сократимость:
Гладкие мышцы способны осуществлять относительно медленные ритмические и длительные тонические сокращения.
Медленные ритмические сокращения обеспечивают перемещение содержимого органа из одной области в другую.
Длительные тонические сокращения, особенно сфинктеров полых органов, препятствуют выходу из них содержимого.
Пластичность:
Это способность сохранять приданную им при растяжении или деформации форму. Благодаря пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии.
Автоматия:
Особенность гладких мышц, отличающая их от скелетных. Благодаря автоматии гладкие мышцы могут сокращаться в условиях отсутствия иннервации. Важную роль в этом играет растяжение.
Растяжение является адекватным раздражителем для гладкой мускулатуры. Сильное и резкое растяжение гладких мышц вызывает их сокращение.
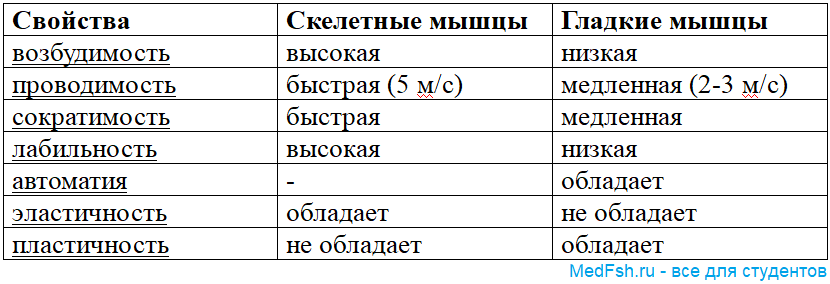
Сравнительная характеристика скелетных и гладких мышц:
Суммирование [ править ]
Каждый нейрон соединяется с множеством других нейронов, получая от них многочисленные импульсы. Суммирование — это сложение этих импульсов на бугорке аксона. Если нейрон получает только возбуждающие импульсы, он генерирует потенциал действия. Если вместо этого нейрон получает столько же тормозных импульсов, сколько и возбуждающих, торможение нейтрализует возбуждение, и нервный импульс на этом останавливается. Генерация потенциала действия пропорциональна вероятности и характеру высвобождения нейромедиаторов, а также постсинаптической сенсибилизации рецепторов.
Пространственное суммирование означает, что эффекты импульсов, полученных в разных местах нейрона, складываются, так что нейрон может срабатывать, когда такие импульсы принимаются одновременно, даже если каждого импульса самого по себе не будет достаточно, чтобы вызвать срабатывание.
Временное суммирование означает, что эффекты импульсов, полученных в одном и том же месте, могут складываться, если импульсы принимаются в близкой временной последовательности. Таким образом, нейрон может срабатывать при получении нескольких импульсов, даже если каждого импульса самого по себе недостаточно, чтобы вызвать срабатывание.
Физиология мышечных волокон
Три типа мышц:
- скелетная (40-50% массы тела),
- сердечная (менее 1%),
- гладкая (8-9%).
Физиологические свойства скелетных мышц:
- Возбудимость — способность отвечать на действие раздражителя возбуждением.
- Проводимость — способность проводить возбуждение из места его возникновения к другим участкам мышцы.
- Лабильность — способность мышцы сокращаться в соответствии с частотой действия раздражителя (200-300 Гц для скелетной мышцы).
- Сократимость — для мышцы является специфическим свойством — это способность мышцы изменять длину или напряжение в ответ на действие раздражителя.
Физические свойства скелетных мышц:
- Растяжимость — способность мышцы изменять длину под действием растягивающей силы.
- Эластичность — способность мышцы восстанавливать первоначальную длину или форму после прекращения действия растягивающей силы.
- Силы мышц — способность мышцы поднять максимальный груз.
- Способность мышцы совершать работу.
Режимы сокращения:
- Изотонический,
- Изометрический,
- Ауксотонический.
Изотонический режим — сокращение мышцы происходит с изменением ее длины без изменения напряжения (тонуса) (напр.: сокращение мышц языка).
Изометрический режим — длина постоянная, увеличивается степень мышечного напряжения (тонуса) (напр.: при поднятии непосильного груза).
Ауксотонический режим — одновременно изменяется длина и напряжение мышцы (характерен для обычных двигательных актов).
Связь с электрическими синапсами [ править ]
Электрический синапс представляет собой электрический проводящая связь между двумя примыкающими нейронами , которая образуются при узком зазоре между пре- и постсинаптическими клетками , известными как щелевые контактами . В щелевых соединениях клетки приближаются друг к другу на расстояние примерно 3,5 нм , а не на расстояние от 20 до 40 нм, которое разделяет клетки в химических синапсах. В отличие от химических синапсов, постсинаптический потенциал в электрических синапсах вызывается не открытием ионных каналов химическими передатчиками, а скорее прямым электрическим взаимодействием между обоими нейронами. Электрические синапсы быстрее химических. Электрические синапсы обнаруживаются по всей нервной системе, в том числе в сетчатке , ретикулярном ядре таламуса , неокортексе и в гиппокампе . В то время как химические синапсы обнаруживаются между возбуждающими и тормозящими нейронами, электрические синапсы чаще всего встречаются между меньшими локальными тормозящими нейронами. Электрические синапсы могут существовать между двумя аксонами, двумя дендритами или между аксоном и дендритом. У некоторых рыб и земноводных электрические синапсы могут быть обнаружены в том же конце химического синапса, что и в клетках Маутнера .
Объемная передача [ править ]
Когда нейротрансмиттер высвобождается в синапсе, он достигает своей максимальной концентрации в узком пространстве синаптической щели, но некоторая его часть обязательно диффундирует, прежде чем реабсорбируется или разрушается. Если он диффундирует прочь, он может активировать рецепторы, расположенные либо в других синапсах, либо на мембране вдали от любого синапса. Внесинаптическая активность нейромедиатора известна как объемная передача . Хорошо известно, что такие эффекты в некоторой степени происходят, но их функциональное значение долгое время оставалось предметом споров.
Недавние исследования показывают, что объемная передача может быть преобладающим способом взаимодействия для некоторых специальных типов нейронов. В коре головного мозга млекопитающих класс нейронов, называемых нейроглиаформными клетками, может подавлять другие близлежащие нейроны коры, высвобождая нейромедиатор ГАМК во внеклеточное пространство. По той же вене, ГАМК, высвобождаемая из нейроглиаформных клеток во внеклеточное пространство, также действует на окружающие астроциты , определяя роль объемной передачи в контроле гомеостаза ионов и нейротрансмиттеров. Примерно 78% бутонов нейроглиаформных клеток не образуют классических синапсов. Это может быть первым убедительным примером химического взаимодействия нейронов там, где нет классических синапсов.
Нервно-мышечное соединение
Во время миотического рефлекса рефлекс сгибания CF пресинаптический элемент встречается с двигательной пластиной мышечного волокна, которое состоит из плазматической мембраны, называемой сарколеммой, которая действует как постсинаптический элемент и содержит несколько сотен миофибрилл
Нервно-мышечное соединение исторически очень важно, поскольку именно наблюдения за разгибателем лягушачьей лапы породили электробиологию, которая быстро повлияла на широкую публику, о чем свидетельствует энтузиазм того времени в отношении электрических явлений.
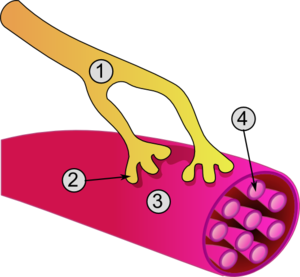 |

|
Детальный вид перекрестка
|
Ацетилхолин участвует в сокращении мышц при сгибании рефлекса или продление на нервно — мышечном соединении. Нейроны, которые его производят, называются холинергическими нейронами. Его предшественниками являются холин пищевого происхождения, который поглощается пресинаптическим концом в крови, и ацетилкофермент А митохондриального происхождения. Они синтезируются ферментом холин-ацетилтрансферазой (CAT), который превращает их в ацетилхолин. Эти нейротрансмиттеры затем покрываются пузырьками, происходящими из зачатка аппарата Гольджи, и транспортируются к синаптическому выступу (или кнопке). На пресинаптическом уровне имеется не одна выпуклость, а сотни, чтобы обеспечить большую контактную поверхность, мы говорим о терминальном ветвлении.
Под действием кальция везикулы, наполненные нейротрансмиттерами, сливаются с плазматической мембраной, выбрасывая свое содержимое в синаптическую щель ( экзоцитоз ). Медиаторы затем прикрепляются к специфическим рецепторам на табличке двигателя в скелетной мышце , которая имеет следствие чего его контракт. Избыток нейротрансмиттер затем разрушаются с помощью фермента : ацетилхолинэстеразы (AChE) , который высвобождает уксусную кислоту и холин , который затем может быть возвращен рецепторами пресинаптического аксона и переработаны.
Синаптический уровень
Структура биологических организмов, составляющих биосферу, может быть разбита на несколько уровней организации : атомный , , клеточный , тканевый , , аппаратные и, наконец, организм в его функциональной целостности.
Научное изучение живых организмов осуществляется путем исследования элементов каждого из этих уровней, а затем путем понимания взаимодействий между этими различными уровнями (см. Статью « »).
Изучение синаптического уровня позволяет понять характеристики передачи биоэлектрических и химических сигналов между самими нейронами или между нейронами и определенными элементами организма ( сенсорными клетками , гладкими или поперечно-полосатыми мышцами и т. Д.). Понимание передачи нейронных сигналов является существенной необходимостью, потому что этот механизм является фундаментальным в функционировании нервной системы.
Механизм сокращения поперечно-полосатых мышц
Любая скелетная мышца состоит из мышечных волокон, которые, в свою очередь, состоят из множества тонких нитей — миофибрилл, расположенных продольно. Каждая миофибрилла состоит из протофибрилл — нитей сократительных белков: миозина (миозиновая протофибрилла), актина (актиновая протофибрилла).
Кроме сократительных белков в миофибрилле имеются два регуляторных белка: тропомиозин и тропонин.
Миозиновые волокна соединены в толстый пучок, от которого в торону актиновых нитей отходят поперечные мостики. У каждого мостика выделяют шейку и головку.
Нить актина располагается в виде 2 скрученных ниток бус. На ней имеются актиновые центры.

Тропомиозин в виде спиралей оплетает поверхность актина, закрывая в покое ее центры. Одна молекула тропомиозина контактирует с 7 молекулами актина.
Тропонин образует утолщение на конце каждой нити тропомиозина.
Под влиянием возникшего в мышечном волокне ПД из саркоплазматического ретикулума (СПР — депо Ca2+) высвобождаются ионы Ca. Кальций связывается с тропонином, который смещает тропомиозиновый стержень, что приводит к открытию актиновых центров.

В результате, к актиновым центрам присоединяются головки поперечных миозиновых мостиков.
Эти постики совершают «гребущие движения», в результате чего нити актина перемещаются этими мостиками относительно волокон миозина, происходит укорочение мышцы.
Процесс расслабления происходит в обратной последовательности с использованием энергии АТФ за счет функционирования кальциевого насоса.
При отсутствии повторного импульса ионы Ca не поступают из СПР. В результате отсутствия Ca-тропонинового комплекса, тропомиозин возвращается на свое прежнее место, блокируя актиновые центры актина. Актиновые протофибриллы легко скользят в обратном направлении благодаря эластичности мышцы, и мышца удлиняется (расслабляется).
Внешние ссылки [ править ]
Послушайте эту статью ( 7 минут )
Этот аудиофайл был создан на основе редакции этой статьи от 19 июня 2005 г. и не отражает последующих правок.
- Обзор Synapse для детей
- Синапсы Biologymad.com (2004)
- Synapse — база данных, ориентированная на ячейки
- Атлас ультраструктурной нейроцитологии Большая картинная галерея синапсов и других нейронных структур, созданная Кристен Харрис в лаборатории Кристен Харрис.
| vтеНервная ткань | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ЦНС |
|
||||||||||||||
| ПНС |
|
||||||||||||||
| Нейроны / нервные волокна |
|
||||||||||||||
| Прекращение |
|
Интеграция сигналов: увеличьте постсинаптический элемент
 Подсознательная деполяризация. Один PPSE не деполяризует мембрану в достаточной степени, чтобы генерировать потенциал действия.
Подсознательная деполяризация. Один PPSE не деполяризует мембрану в достаточной степени, чтобы генерировать потенциал действия.
 Постсинаптическая регистрация мембранного потенциала. Стрелками отмечены PPSE трех связанных событий. Суммирование трех PPSE в этом случае приводит к срабатыванию потенциала действия.
Постсинаптическая регистрация мембранного потенциала. Стрелками отмечены PPSE трех связанных событий. Суммирование трех PPSE в этом случае приводит к срабатыванию потенциала действия.
Синапсы делятся на две категории в зависимости от вызываемого ими эффекта: возбуждающего или тормозящего. Основным тормозным нейромедиатором в головном мозге является ГАМК, которая связывается с каналами рецептора ГАМК А , открытие которых вызывает приток ионов хлора и, следовательно, гиперполяризацию мембраны. Существует большее разнообразие возбуждающих ионотропных рецепторов, например рецепторов глутамата или ацетилхолина . Постсинаптический элемент обычно имеет эти две категории рецепторов, а также натриевые или кальциевые каналы, активируемые деполяризацией. Он выполняет временное суммирование возбуждающих (PPSE, возбуждающий постсинаптический потенциал) и тормозных (PPSI, тормозных постсинаптических потенциалов) сигналов. Он будет распространять потенциал действия при условии, что сумма возбуждений больше, чем сумма запретов, и если будет достигнут порог деполяризации. Этот порог соответствует напряжению, при котором активируется достаточное количество натриевых каналов.
Пространственное суммирование относится к различным синапсам , связанным с постсинаптическим элементом. Нейрон действительно может воспринимать более тысячи различных воздействий, но он может реагировать только одним способом: проводимостью или отсутствием проводимости. Если результат алгебраической суммы всех афферентных элементов превышает пороговое значение, около -15 мВ на диаграмме напротив, интегрирующий нейрон будет местом расположения потенциала действия.
Так называемое временное суммирование также имеет место на уровне постсинаптического элемента. Это связано со скоростью проникновения ионов внутрь клетки. Если вовремя собрать много PPSE, они складываются и также могут достичь порога деполяризации и вызвать потенциал действия.
Заключительный элемент интеграции связан с существованием рефрактерного периода нейрона. Если два афферентных возбуждающих сигнала разнесены менее чем на миллисекунду, второй не вызовет никакого PPSE и, следовательно, будет беззвучным.
Действие наркотиков [ править ]
Одна из наиболее важных особенностей химических синапсов заключается в том, что они являются местом действия большинства психоактивных веществ . На синапсы влияют такие наркотики, как кураре, стрихнин, кокаин, морфин, алкоголь, ЛСД и многие другие. Эти препараты по-разному влияют на синаптическую функцию и часто ограничиваются синапсами, в которых используется конкретный нейромедиатор. Например, кураре — это яд, не позволяющий ацетилхолину деполяризовать постсинаптическую мембрану, вызывая паралич . Стрихнин блокирует тормозящие эффекты нейротрансмиттера глицина , который заставляет организм улавливать и реагировать на более слабые и ранее игнорируемые раздражители, что приводит к неконтролируемым мышечным спазмам.. Морфин действует на синапсы, в которых используются нейромедиаторы эндорфина , а алкоголь усиливает тормозящие эффекты нейромедиатора ГАМК . ЛСД влияет на синапсы, которые используют нейромедиатор серотонин . Кокаин блокирует обратный захват дофамина и, следовательно, усиливает его действие.
Особенности проведения возбуждения в нервно-мышечном синапсе
Одностороннее проведение возбуждения — только в направлении от пресинаптического окончания к постсинаптической мембране.
Суммация возбуждения соседних постсинаптических мембран.
Синаптическая задержка — замедление в проведении импульса от нейрона к мышце составляет 0,5-1 мс. Это время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, взаимодействие с рецептором, формирование ПКП, их суммацию.
Низкая лабильность — она составляет 100-150 имп/с для сигнала, что в 5-6 раз ниже лабильности нервного волокна.
Чувствительность к действию лекарственных веществ, ядов, БАВ, выполняющих роль медиатора.
Утомляемость химических синапсов — выражается в ухудшении проводимости вплоть до блокады в синапсе при длительном функционировании синапса. Главная причина утомляемости — исчерпание запасов медиатора в пресинаптическом окончании.
Законы проведения возбуждения по нервам:
- Закон функциональной целостности нерва.
- Закон изолированного проведения возбуждения.
- Закон двустороннего проведения возбуждения.
В зависимости от скорости проведения возбуждения нервные волокна подразделяются на 3 группы: A, B, C. В группе A выделяют 4 подгруппы: альфа, бетта, гамма и сигма.
Механизм синаптической передачи
4 этап
Ионы Ca вызывают образование специального белкового комплекса, который включает в себя везикулу и структуры, расположенные непосредственно около пресинаптической мембраны.
Они связаны между собой так называемыми белками экзоцитоза.
Часть белков расположена на везикулах (синапсин, синаптотагмин, синаптобревин), а часть — на пресинаптической мембране (синтаксин, синапсо-ассоциированный белок). Данный комплекс получил название секретосома.

Излитию содержимого пузырька в щель способствует белок синаптопорин, формирующий канал, по которому идет выброс медиатора.
Квант медиатора — количество молекул, содержащихся в одной везикуле.
На 1 ПД выбрасывается 100 квантов АХ.
10 этап

На постсинаптической мембране возникает потенциал концевой пластинки (ПКП). Он является аналогом локального ответа (ЛО).
Потенциал действия на постсинаптической мембране не возникает! Он формируется на соседней мембране мышечного волокна.
Судьба медиатора:
- связывание с рецептором,
- разрушение ферментов (ацетилхолинэстеразой),
- обратное поглощение в пресинаптическую мембрану,
- вымывание из щели и фагоцитоз.
События в синапсе:
- ПД приходит к терминали аксона;
- Он деполяризует пресинаптическую мембрану;
- Ca2+ входит в терминаль, что приводит к выделению АХ;
- В синаптическую щель выделяется медиатор АХ;
- Он диффундирует в щель и связывается с рецепторами постсинаптической мембраны;
- Меняется проницаемость постсинаптической мембраны для ионов Na+;
- Ионы Na+ проникают в постсинаптическую мембрану и уменьшают ее заряд — возникает потенциал концевой пластинки (ПКП).
На самой постсинаптической мембране ПД возникнуть не может, так как здесь отсутствуют потенциалзависимые каналы, они являются хемозависимыми!
- ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и его распространению по мышечному волокну (около 5 м/с).
Достигнув пороговой величины, то есть КУД, ПКП возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна за счет местных круговых токов.

Cotransmission [ править ]
Котрансмиссия — это высвобождение нескольких типов нейромедиаторов из одного нервного окончания .
На нервном окончании нейротрансмиттеры присутствуют в покрытых мембраной пузырьках размером 35-50 нм, называемых синаптическими пузырьками . Для высвобождения нейротрансмиттеров синаптические везикулы временно стыкуются и сливаются в основании специализированных чашеобразных липопротеидных структур 10–15 нм на пресинаптической мембране, называемых поросомами . Протеом нейрональных поросом был решен, обеспечивая молекулярную архитектуру и полную композицию механизма.
Недавние исследования множества систем показали, что большинство, если не все, нейроны испускают несколько различных химических посредников. Котрансмиссия допускает более сложные эффекты на постсинаптические рецепторы и, таким образом, обеспечивает более сложную коммуникацию между нейронами.
В современной нейробиологии нейроны часто классифицируют по их котрансмиттеру. Например, «ГАМКергические нейроны» полосатого тела используют опиоидные пептиды или вещество P в качестве своего основного котрансмиттера.
Некоторые нейроны могут высвобождать по крайней мере два нейротрансмиттера одновременно, а другой является котрансмиттером, чтобы обеспечить стабилизирующую отрицательную обратную связь, необходимую для значимого кодирования, в отсутствие тормозящих интернейронов . Примеры включают:
- ГАМК — совместный выпуск глицина .
- Дофамин — глутамат со-релиз.
- Совместное высвобождение ацетилхолина (Ach) и глутамата.
- Совместное высвобождение ACh– вазоактивного кишечного пептида (VIP).
- Совместное высвобождение ACh -пептида, связанного с геном кальцитонина (CGRP).
- Совместное высвобождение глутамата и динорфина (в гиппокампе ).
Норадреналин и АТФ являются симпатическими передатчиками. Установлено , что эндоканнабиноид анандамид и каннабиноиды , WIN 55,212-, 2 могут изменить общую реакцию на симпатическую стимуляцию нерва, и указует , что пресинаптические рецепторы СВ1 опосредовать симпато -inhibitory действия. Таким образом, каннабиноиды могут подавлять как норадренергические, так и пуринергические компоненты симпатической нейротрансмиссии.
Одной из необычных пар ко-трансмиттеров является ГАМК и глутамат, которые выделяются из одних и тех же окончаний аксонов нейронов, происходящих из вентральной тегментальной области (VTA), внутреннего бледного шара и супраммиллярного ядра . Первые два проецируются на габенулу, тогда как выступы супраммиллярного ядра, как известно, нацелены на зубчатую извилину гиппокампа.






