Энергетический обмен в егэ по биологии
Содержание:
- Физиологическое значение
- Световая фаза фотосинтеза
- Этапы окислительного гликолиза. Фаза 1.
- Обходной путь 1. Образование фосфоенолпирувата
- Что такое энергетический обмен?
- Обходной путь 3. Образование глюкозы
- Что нужно запомнить?
- Общая характеристика систем энергообеспечения мышечной деятельности.
- Примеры заданий
- Бескислородное окисление глюкозы включает два этапа
- Путь[править]
- Что такое гликолиз
Физиологическое значение
Гликолиз можно считать универсальным способом катаболизма глюкозы. Его активно используют про- и эукариотические организмы. Ферменты, являющиеся катализаторами гликолиза, растворяются в воде, накапливаются в цитозоле. Часть клеток и тканей животных могут катаболизировать гексозу только путем гликолиза. Например, подобные способности есть у клеток почечных каналов, нейронов мозга.
В жировой ткани и печени есть некоторые отличия в физиологической роли гликолиза. При пищеварении в жировой ткани и в печени данный процесс является источником субстратов, используемых в синтезе жиров.
Часть тканей растений запасают крахмал в клубнях. Водные растения энергию получают через гликолиз.
В анаэробных условиях происходит превращение пирувата в этанол и лактат. Процесс сопровождается выделением большого количества энергии.
Гликолиз имеет существенное физиологическое значение в адипоцитах. С его помощью вместо окислительного процесса происходит липогенез, позволяющий снижать окислительный стресс.
Световая фаза фотосинтеза
Чтобы лучше понять, что происходит во время фотосинтеза, разберём фазы фотосинтеза. Световая фаза фотосинтеза включает в себя фотохимические и фотофизические процессы, и может быть поделена на три этапа:
- Фаза поглощения — энергия света улавливается при помощи светособирающих комплексов, переходит в энергию электронного возбуждения пигментов, передаётся в реакционный центр фотосистем I и II.
- Фаза реакционных центров — энергия электронного возбуждения пигментов светособирающих комплексов используется для активации реакционных центров фотосистем. В реакционном центре электрон от возбуждённого хлорофилла передаётся другим компонентам электрон-транспортной цепи, пигмент после отдачи электрона переходит в окисленное состояние и становится способным, в свою очередь, отнимать электроны у других веществ. Именно в этом процессе происходит преобразование физической формы энергии в химическую.
- Фаза электрон-транспортной цепи — электроны переносятся по цепи переносчиков, образуются АТФ, НАДФН, O2. Необходимо, чтобы каждый переносчик электрон-транспортной цепи поочерёдно восстанавливался и окислялся, обеспечивая таким образом перенос энергии электронов. Любой этап переноса электрона сопровождается высвобождением или поглощением энергии. Часть энергии теряется. На некоторых участках электрон-транспортной цепи перенос электрона сопряжён с переносом протона.
Для того чтобы понять, что происходит во время фазы фотосинтеза, рассмотрим эти процессы подробнее. Кванты света улавливаются светособирающими комплексами фотосистемы I — молекула хлорофилла в составе светособирающего комплекса переходит в возбуждённое состояние, и энергия передаётся в реакционный центр фотосистемы I. Происходит возбуждение молекул хлорофилла фотосистемы I, отщепляется электрон. Пройдя по цепочке внутренних компонентов фотосистемы I и внешних переносчиков, электрон в конце концов попадает к НАДФ+ — образуется восстановитель НАДФН. Получается, что хлорофилл фотосистемы I отдал электрон и приобрёл положительный заряд, и для дальнейшего функционирования необходимо восстановить нейтральность молекулы, получить электрон, чтобы закрыть «дырку». Этот электрон приходит от фотосистемы II.
На светособирающие комплексы фотосистемы II попадают кванты света — происходит возбуждение молекулы хлорофилла фотосистемы II, молекула хлорофилла отдаёт электрон и переходит в окисленное состояние. Нехватку электрона хлорофилл восполняет благодаря фотолизу воды, при этом образуется протоны H+, а также важный побочный продукт фотосинтеза — кислород. По цепи переносчиков электрон от хлорофилла фотосистемы II попадает к хлорофиллу реакционного центра фотосистемы I и восстанавливает его. Теперь этот хлорофилл может снова поглощать энергию кванта света и отдавать электрон в электрон-транспортную цепь.
Протоны, попадающие во внутритилакоидное пространство, используются для синтеза АТФ. С помощью фермента АТФ-синтазы за счёт градиента протонов образуется АТФ из АДФ и фосфата. Под градиентом понимают неравномерное распределение: во внутритилакоидном пространстве H+ больше, в строме — меньше. Поэтому частицы стремятся проникнуть в строму, переходят в неё через АТФ-синтазу, а в процессе пути сквозь белковый комплекс отдают ему часть энергии, которая и используется для синтеза АТФ.
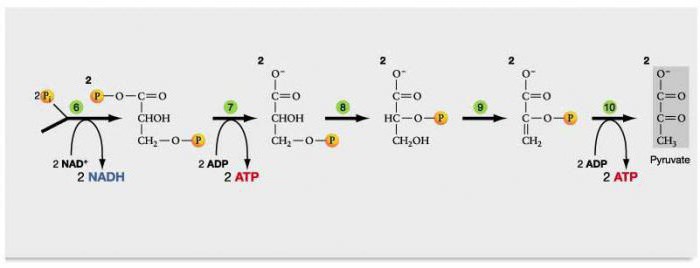
Этапы окислительного гликолиза. Фаза 1.
В аэробном гликолизе выделяются 2 фазы.
Первая фаза – подготовительная. В ней глюкоза вступает в реакцию с 2 молекулами АТФ. Эта фаза состоит из 5 последовательных ступеней биохимических реакций.
1-я ступень. Фосфорилирование глюкозы
Фосфорилирование, то есть процесс переноса остатков фосфорной кислоты в первой и последующих реакциях производится за счет молекул адезинтрифосфорной кислоты.
В первой ступени остатки фосфорной кислоты из молекул адезинтрифосфата переносятся в молекулярную структуру глюкозы. В ходе процесса получается глюкозо-6-фосфат. В качестве катализатора в процессе выступает гексокиназа, ускоряющая процесс с помощью ионов магния, выступающих в качестве кофактора. Ионы магния задействованы и в других реакциях гликолиза.

2-я ступень. Образование изомера глюкозо-6-фосфата

На 2-й ступени происходит изомеризация глюкозо-6-фосфата во фруктозу-6-фосфат.
Изомеризация – образование веществ, имеющих одинаковый вес, состав химических элементов, но обладающих разными свойствами вследствие различного расположения атомов в молекуле. Изомеризация веществ осуществляется под действием внешних условий: давления, температур, катализаторов.
В данном случае процесс осуществляется под действием катализатора фосфоглюкозоизомеразы при участии ионов Mg+.
3-я ступень. Фосфорилирование фруктозо-6-фосфата
На данной ступени происходит присоединение фосфорильной группы за счет АТФ. Процесс осуществляется при участии фермента фосфофруктокиназа-1. Этот фермент и предназначен только для участия в гидролизе. В результате реакции получаются фруктозо-1,6-бисфосфат и нуклеотид адезинтрифосфат.

АТФ – адезинтрифосфат, уникальный источник энергии в живом организме. Представляет собой довольно сложную и громоздкую молекулу, состоящую из углеводородных, гидроксильных групп, азота и групп фосфорной кислоты с одной свободной связью, собранных в нескольких циклических и линейных структурах. Высвобождение энергии происходит в результате взаимодействия остатков фосфорной кислоты с водой. Гидролиз АТФ сопровождается образованием фосфорной кислоты и выделением 40-60 Дж энергии, которую организм затрачивает на свою жизнедеятельность.
Но прежде должно произойти фосфорилирование глюкозы за счет молекулы Адезинтрифосфата, то есть перенос остатка фосфорной кислоты в глюкозу.
4-я ступень. Распад фруктозо-1,6-дифосфата

В четвертой реакции фруктозо-1,6-дифосфат распадается на два новых вещества.
- Диоксиацетонфосфат,
- Глицеральд альдегид-3-фосфат.
В данном химическом процессе в качестве катализатора выступает альдолаза, фермент, участвующий в энергетическом обмене, и необходимый при диагностировании ряда заболеваний.
5-я ступень. Образование триозофосфатных изомеров
И, наконец, последний процесс – изомеризация триозофосфатов.

Глицеральд-3-фосфат продолжит участвовать в процессе аэробного гидролиза. А второй компонент – диоксиацетон фосфат при участии фермента триозофосфатизомеразы преобразуется в глицеральдегид-3-фосфат. Но трансформация эта – обратимая.
Обходной путь 1. Образование фосфоенолпирувата
Пируват, который образовался в ходе гликолиза был транспортирован в митохондрию. Нам повезло, потому что первый фермент обходного пути тоже находится в матриксе митохондрии. Этот фермент — митохондриальная пируваткарбокилаза.
Пируваткарбоксилаза присоединяет к пирувату CO2 — образуется оксалоацетат.
Первая реакция в образовании фосфоенолпирувата. Идёт в матриксе митохондрий.
Для этой реакции нужны молекула АТФ и кофермент — биотин. АТФ тратится на активацию бикарбонатного иона, так как у нас в организме большая часть CO2 растворена. Из бикарбонатного иона образуется CO2, который присоединяется к биотину. Биотин переносит CO2 на молекулу пирувата, клетка получает оксалоацетат.
Дальше нужно перенести оксалоацетат в цитоплазму клетки — там находятся семь ферментов, которые катализируют обратимые реакции гликолиза. Для оксалоацетата во внутренней мембране митохондрий нет переносчика. Но есть для малата. Поэтому сначала оксалоацетат превращается в малат. Катализатор — митохондриальная малатдегидрогеназа. Она присоединяет два протона и два электрона к молекуле оксалоацетата. Кофермент реакции — НАДH. Так образуется малат.
Образование малата в митохондрии
Митохондриальная малатдегидрогеназа обычно катализирует обратную реакцию — превращение малата в оксалоацетат. Так было в но почему здесь всё по-другому? Дело в том, что в митохондрии находится избыток НАДH, а не НАД+. Поэтому реакция идёт в другую сторону. Избыток НАДH обеспечивает окисление жирных кислот — запускается при голодании и/или физической нагрузке.
Далее малат транспортируется через внутреннюю мембрану с помощью белка-переносчика. В наружной мембране есть поры, поэтому дальше он влетает в цитоплазму.
Транспорт малата в цитоплазму
В цитоплазме клетки малат превращается в оксалоацетат с помощью цитоплазматической малатдегидрогеназы. При этом образуется оксалоацетат и НАДH. Эта реакция полезна, так как в шестой реакции для образования глицеральдегид-3-фосфата нужен НАДH. Так митохондрия передаёт избыток протонов и электронов на нужды цитоплазмы — всё это через образование малата.
Образование оксалоацетата в цитоплазме клетки
Теперь нужно превратить оксалоацетат в фосфоенолпируват. Для этого нужен второй фермент этого обходного пути — фосфоенолпируваткарбоксикиназа. Он получил название по обратной реакции. Кроме фермента, нужен источник энергии и фосфатной группы — здесь используется ГТФ. Образуется фосфоенолпируват, ГДФ и CO2. Так клетка обходит десятую реакцию гликолиза.
Вторая реакция обходного пути. Образование фосфоенолпирувата с помощью декарбоксилирования оксалоацетата
Пока что глюконеогенез дорогой процесс. Мы затратили молекулу АТФ и ГТФ на образование фосфоенолпирувата. А эта молекула содержит только три атома углерода. Для образования глюкозы нужно образовать два фосфоенолпирувата. То есть общие затраты энергии — 2 молекулы АТФ и 2 ГТФ.
Дальше идут обратимые реакции до образования фруктозо-1,6-бисфосфата (третья реакция гликолиза). В ходе седьмой реакции мы затратим ещё две молекулы АТФ на образование двух 1,3-бисфосфоглицератов.
Общие затраты энергии — 4 молекулы АТФ и 2 молекулы ГТФ. Дальше клетка не будет тратиться на синтез глюкозы, так что это общие затраты на весь глюконеогенез.
Хоть этот процесс и дорогой, но других вариантов у организма нет. Нельзя же бросить мозг и эритроциты — без них тело долго не проживёт.
Что такое энергетический обмен?
Для начала нужно разобраться, что такое энергетический обмен и какие у него есть особенности. Уверена, что вы встречали в тестах слова «катаболизм» и «диссимиляция», эти названия являются синонимами термина «энергетический обмен», советую их запомнить. Что же такое энергетический обмен? Это реакции, при которых органические вещества расщепляются, а энергия запасается клеткой в молекулах АТФ. Эту энергию клетка потом потратит на дальнейшую жизнедеятельность.
Такой тип обмена (как и все реакции метаболизма) идет поэтапно. В нем выделяют два или три основных этапа — это зависит от организации клетки и среды, в которой она обитает. Предлагаю рассмотреть каждый из этапов энергетического обмена подробнее.
Если хотите лучше понять не только энергетический обмен, но и другие темы ЕГЭ по биологии, приходите учиться в MAXIMUM! Записывайтесь на консультацию — вы сможете пройти диагностику по выбранным предметам ЕГЭ, поставить цели и составить стратегию подготовки, чтобы получить на экзамене высокие баллы. Все это абсолютно бесплатно!
Обходной путь 3. Образование глюкозы
От глюкозо-6-фосфата отщепляется фосфатная группа. Катализирует реакцию глюкозо-6-фосфатаза.
Образование глюкозы в третьем обходном пути
Но этот фермент не может находиться в цитоплазме. Почему? Тогда клетка не смогла бы выполнять свои функции. Представьте, что глюкоза попадает в печень. Она сразу же фосфорилируется с помощью глюкокиназы. Так клетка добивается двух вещей:
- Увеличивает реакционную способность глюкозы;
- Не даёт глюкозе выбраться из клетки. Потому что для глюкозо-6-фосфата нет белков переносчиков в цитоплазме. А вот для обычной глюкозы есть —
Если мы засунем в эту систему фермент глюкозо-6-фосфатазу, то она будет постоянно превращать глюкозо-6-фосфат в глюкозу. А та будет вылетать из клетки. Получается, что вся система работает в холостую. Поэтому глюкозо-6-фосфатаза не работает в цитоплазме, а находится в эндоплазматическом ретикулуме (ЭПР). Он отделён от цитоплазмы биологической мембраной.
В мембране ЭПР есть белки переносчики для глюкозо-6-фосфата. Он переносится в ЭПР, где дефосфорилируется глюкозо-6-фосфатазой. Образуется глюкоза и неорганический фосфат. Они переносятся в цитоплазму, опять же с помощью своих белков переносчиков. А дальше глюкоза попадает в кровь через глюкозный транспортёр второго типа.
Глюкозо-6-фосфатаза работает в эндоплазматическом ретикулуме. Так клетка избегает холостых реакций.
Вот и весь глюконеогенез. Держите общую картинку.
Пируваткарбоксилаза не влезла, извиняйте.
Что нужно запомнить?
- Энергетический обмен нужен, чтобы запасать энергию, расщепляя полимеры
- Первый этап энергетического обмена — подготовительный. В пищеварительной системе и /или лизосомах, полимеры распадаются до мономеров, энергия расходуется в виде тепла
- Второй этап — гликолиз. Проиходит в цитоплазме. Глюкоза распадается до 2 молекул ПВК, запасается 2 АТФ
- Третий этап — клеточное дыхание. Происходит на кристах митохондрий. ПВК полностью окисляется, запасается 36 молекул АТФ
- За три этапа клетка может получить 38 АТФ из одной молекулы глюкозы: 2 АТФ на втором и 36 АТФ на третьем
ЕГЭ по биологии — большой и сложный экзамен, который состоит из большого количества тем и заданий. Но сдать его на высокий балл реально, если организовать систематическую подготовку. Обязательно приходите на бесплатную консультацию в MAXIMUM — там вы сможете построить индивидуальную стратегию подготовки к ЕГЭ и узнаете все подводные камни экзамена.
Общая характеристика систем энергообеспечения мышечной деятельности.
Энергия, как известно, представляет собой общую количественную меру, связывающую воедино все явления природы, разные формы движения материи
Из всех видов энергии, образующейся и использующейся в различных физических процессах(тепловая, механическая, химическая и др.)применительно к мышечной деятельности, основное внимание должно быть сконцентрировано на химической энергии организма, источником которой являются пищевые продукты и её преобразовании в механическую энергию двигательной деятельности человека
Энергия, высвобождаемая во время расщепления пищевых продуктов, используется для производства аденозинтрифосфата (АТФ), который депонируется в мышечных клетках и является своеобразным топливом для производства механической энергии мышечного сокращения.
Энергию для мышечного сокращения даёт расщепление аденозинтрифосфата (АТФ) до аденозиндифосфата (АДФ) и неорганического фосфата (Ф). Количество АТФ в мышцах невелико и его достаточно для обеспечения высокоинтенсивной работы лишь в течении 1 – 2 с. Для продолжения работы необходим ресинтез АТФ, который производится за счет энерго отдающих реакций трёх типов. Восполнение запасов АТФ в мышцах позволяет поддерживать постоянный уровень его концентрации, необходимый для полноценного мышечного сокращения.
Ресинтез АТФ обеспечивается как в анаэробных, так и в аэробных реакциях с привлечением в качестве энергетических источников запасов креатинфосфата (КФ) и АДФ, содержащихся в мышечных тканях, а также богатых энергией субстратов (гликоген мышц и печени, запасы липозной ткани и др.). Химические реакции, приводящие к обеспечению мышц энергией протекают в трёх энергетических системах:
- анаэробной алактатной
- анаэробной лактатной (гликолитической)
- аэробной.
Образование энергии в первых двух системах осуществляется в процессе химических реакций, не требующих наличия кислорода. Третья система предусматривает энергообеспечение мышечной деятельности в результате реакций окисления, протекающих с участием кислорода. Наиболее общие представления о последовательности включения и количественных соотношениях в энергообеспечении мышечной деятельности каждой из указанных систем приведены на рис. 1.

Рис. 1. Последовательность и количественные соотношения процессов энергообеспечения мышечной деятельности у квалифицированных спортсменов в различных энергетических системах (схема): 1 – алактатной; 2 – лактатной; 3 – аэробной.
Возможности каждой из указанных энергетических систем определяются мощностью, т. е. скоростью освобождения энергии в метаболических процессах, и ёмкостью, которая определяется величиной и эффективностью использования субстратных фондов.
Примеры заданий
Давайте разберем несколько заданий на энергетический обмен из ЕГЭ по биологии, чтобы закрепить знания на практике.
Пример 1. Что характерно для аэробного этапа энергетического процесса?
- протекает в лизосомах
- расщепляются молекулы ПВК
- наблюдается высокий выход молекул АТФ
- проходит в цитоплазме
- встречается у бактерий
- имеются циклические реакции
Решение. Аэробный или кислородный этап — третий этап энергетического обмена. Он проходит на кристах митохондрий, там расположены ферментативные комплексы и идут циклические реакции, в которых молекулы пировиноградной кислоты разрушаются, на этом этапе наблюдается высокий выход энергии —36 АТФ. В лизосомах проходит подготовительный этап, а в цитоплазме — гликолиз. Кислородный этап не характерен для бактерий, так как у них нет мембранных органоидов.
Ответ: 236
Пример 2. Установите соответствие между характеристикой энергетического обмена и его этапом
| ХАРАКТЕРИСТИКА | ЭТАП ЭНЕРГЕТИЧЕСКОГО ОБМЕНА | |
| A) происходит в аэробных условияхБ) происходит в цитоплазмеB) образуется молочная кислотаГ) образуется пировиноградная кислотаД) синтезируется 36 молекул АТФ | 1) гликолиз2) кислородное окисление |
Решение. Гликолиз — второй этап энергетического обмена, анаэробный, проходит в цитоплазме, образуется пировиноградная кислота, а при недостатке кислорода еще и молочная кислота. Кислородный — третий этап, аэробный, завершается образованием 36 молекул АТФ.
Ответ: 21112
Пример 3. В процессе гликолиза образовались 64 молекулы пировиноградной кислоты (ПВК). Какое количество молекул глюкозы подверглось расщеплению и сколько молекул АТФ образуется при полном окислении глюкозы в клетках эукариот? Ответ поясните
Решение:
- Во время гликолиза одна молекула глюкозы распадается до двух молекул пировиноградной кислоты. Для образования 64 молекул ПВК расщепилось 32 молекулы глюкозы (64:2).
- При полном окислении одной молекулы глюкозы в эукариотической клетке образуется 38 молекул АТФ. При расщеплении 32 молекул глюкозы образуется 1216 молекул АТФ (38*32).
Как видите, энергетический обмен — важная часть ЕГЭ по биологии. Справиться с заданиями достаточно просто, если знать, что происходит на каждом из этапов.
Бескислородное окисление глюкозы включает два этапа
В анаэробном процессе пировиноградная кислота восстанавливается до молочной кислоты (лактата), поэтому в микробиологии анаэробный гликолиз называют молочнокислым брожением. Лактат является метаболическим тупиком и далее ни во что не превращается, единственная возможность утилизовать лактат – это окислить его обратно в пируват.
Многие клетки организма способны к анаэробному окислению глюкозы. Для эритроцитов он является единственным источником энергии.
Клетки скелетной мускулатуры за счет бескислородного расщепления глюкозы способны выполнять мощную, быструю, интенсивную работу, как, например, бег на короткие дистанции, напряжение в силовых видах спорта.
Вне физических нагрузок бескислородное окисление глюкозы в клетках усиливается при гипоксии – при различного рода анемиях, при нарушении кровообращения в тканях независимо от причины.
Первый этап гликолиза – подготовительный, здесь происходит затрата энергии АТФ, активация глюкозы и образование из нее триозофосфатов.
Первая реакция гликолиза сводится к превращению глюкозы в реакционно-способное соединение за счет фосфорилирования 6-го, не включенного в кольцо, атома углерода. Эта реакция является первой в любом превращении глюкозы, катализируется гексокиназой.
Вторая реакция необходима для выведения еще одного атома углерода из кольца для его последующего фосфорилирования (фермент глюкозофосфат-изомераза). В результате образуется фруктозо-6-фосфат.
Третья реакция – фермент фосфофруктокиназа фосфорилирует фруктозо-6-фосфат с образованием почти симметричной молекулы фруктозо-1,6-дифосфата. Эта реакция является главной в регуляции скорости гликолиза.
В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат-альдолазой с образованием двух фосфорилированных триоз-изомеров – альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ).
Пятая реакция подготовительного этапа – переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга при участии триозофосфатизомеразы. Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97%, доля глицеральдегидфосфата – 3%. Эта реакция, при всей ее простоте, определяет дальнейшую судьбу глюкозы:
- при нехватке энергии в клетке и активации окисления глюкозы диоксиацетонфосфат превращается в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза,
- при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний отправляется на синтез жиров.
Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ.
Шестая реакция гликолиза (фермент глицеральдегидфосфат-дегидрогеназа) – окисление глицеральдегидфосфата и присоединение к нему фосфорной кислоты приводит к образованию макроэргического соединения 1,3-дифосфоглицериновой кислоты и НАДН.
В седьмой реакции (фермент фосфоглицераткиназа) энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тратится на образование АТФ.
Реакция получила дополнительное название – реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (от субстрата реакции) в отличие от окислительного фосфорилирования (от электрохимического градиента ионов водорода на мембране митохондрий).
Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы изомеризуется в 2-фосфоглицерат.
Девятая реакция – фермент енолаза отрывает молекулу воды от 2-фосфоглицериновой кислоты и приводит к образованию макроэргической фосфоэфирной связи в составе фосфоенолпирувата.
Десятая реакция гликолиза – еще одна реакция субстратного фосфорилирования – заключается в переносе пируваткиназой макроэргического фосфата с фосфоенолпирувата на АДФ и образовании пировиноградной кислоты.
Последняя реакция бескислородного окисления глюкозы, одиннадцатая – образование молочной кислоты из пирувата под действием лактатдегидрогеназы
Важно то, что эта реакция осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, образующийся в 6-й реакции, в отсутствие кислорода не может окисляться в митохондриях
| У плода и детей первых месяцев жизни преобладает анаэробный распад глюкозы, в связи с чем концентрация молочной кислоты в крови у них выше чем у взрослых. |
| При наличии кислорода пировиноградная кислота переходит в митохондрию и превращается в ацетил-S-КоА. |
Путь[править]
Первой реакцией гликолиза является фосфорилирование молекулы глюкозы, происходящее при участии тканеспецефичного фермента гексокиназы с затратой энергии 1 молекулы АТФ; образуется активная форма глюкозы — глюкозо-6-фосфат (Г-6-Ф):
Для протекания реакции необходимо наличие в среде ионов Mg2+, с которым комплексно связывается молекула АТФ. Эта реакция необратима и является первой ключевой реакцией гликолиза.
Фосфорилирование глюкозы преследует две цели: во-первых, из-за того что плазматическая мембрана, проницаемая для нейтральной молекулы глюкозы, не пропускает отрицательно заряженные молекулы Г-6-Ф, фосфорилированная глюкоза оказывается запертой внутри клетки. Во-вторых, при фосфорилировании глюкоза переводится в активную форму, способную участвовать в биохимических реакциях и включаться в метаболические циклы. Фосфорилирование глюкозы — это единственная реакция в организме, в которой глюкоза участвует как таковая.
Печёночный изофермент гексокиназы — глюкокиназа — имеет важное значение в регуляции уровня глюкозы в крови.
В следующей реакции (2) ферментом фосфоглюкоизомеразой Г-6-Ф превращается во фруктозо-6-фосфат (Ф-6-Ф):
Энергия для этой реакции не требуется, и реакция является полностью обратимой. На данном этапе в процесс гликолиза может также включаться путём фосфорилирования и фруктоза.
Далее почти сразу друг за другом следуют две реакции: необратимое фосфорилирование фруктозо-6-фосфата (3) и обратимое альдольное расщепление образовавшегося фруктозо-1,6-бифосфата (Ф-1,6-бФ) на две триозы (4).
Фосфорилирование Ф-6-Ф осуществляется фосфофруктокиназой с затратой энергии ещё одной молекулы АТФ; это вторая ключевая реакция гликолиза, её регуляция определяет интенсивность гликолиза в целом.
Альдольное расщепление Ф-1,6-бФ происходит под действием альдолазы фруктозо-1,6-бифосфата:
В результате четвёртой реакции образуются дигидроксиацетонфосфат и глицеральдегид-3-фосфат, причём первый почти сразу под действием фосфотриозоизомеразы переходит во второй (5), который и участвует в дальнейших превращениях:
Каждая молекула глицеральдегидфосфата окисляется НАД+ в присутствии дегидрогеназы глицеральдегидфосфата до 1,3-дифосфоглицерата (6):
Далее с 1,3-дифосфоглицерата, содержащего макроэргическую связь в 1 положении, ферментом фосфоглицераткиназой на молекулу АДФ переносится остаток фосфорной кислоты (реакция 7) — образуется молекула АТФ:
Это первая реакция субстратного фосфорилирования. С этого момента процесс расщепления глюкозы перестаёт быть убыточным в энергетическом плане, так как энергетические затраты первого этапа оказываются компенсированными: синтезируются 2 молекулы АТФ (по одной на каждый 1,3-дифосфоглицерат) вместо двух потраченных в реакциях 1 и 3. Для протекания данной реакции требуется присутствие в цитозоле АДФ, то есть при избытке в клетке АТФ (и недостатке АДФ) её скорость снижается. Поскольку АТФ, не подвергающийся метаболизму, в клетке не депонируется а просто разрушается, то эта реакция является важным регулятором гликолиза.
Затем последовательно: фосфоглицеролмутаза образует 2-фосфоглицерат (8):
Енолаза образует фосфоенолпируват (9):
И наконец происходит вторая реакция субстратного фосфорилирования АДФ с образованием енольной формы пирувата и АТФ (10):
Реакция протекает под действием пируваткиназы. Это последняя ключевая реакция гликолиза. Изомеризация енольной формы пирувата в пируват происходит неферментативно.
С момента образования Ф-1,6-бФ с выделением энергии протекают только реакции 7 и 10, в которых и происходит к субстратное фосфорилировнаие АДФ.
Что такое гликолиз
В общих чертах, гликолиз — это процесс, который происходит в цитоплазме всех клеток: от низших бактерий до высших млекопитающих — людей. Для некоторых это единственный метаболический путь доставки энергии клетке — например, для бактерий, для других же это метаболический путь, в который «сливаются» и другие метаболиты — белки и липиды.
Гликолиз — это основной путь расщепления глюкозы в нашем организме, основное биологическое значениеэтого процесса заключается в получении энергии и других продуктов обмена.
 Гликолиз происходит как в аэробных, так и в анаэробных условиях. Процесс в анаэробных условиях (недостаток кислорода) называется анаэробным гликолизом — конечным продуктом лактата и ферментации в дрожжах — алкогольной или молочнокислой ферментации.
Гликолиз происходит как в аэробных, так и в анаэробных условиях. Процесс в анаэробных условиях (недостаток кислорода) называется анаэробным гликолизом — конечным продуктом лактата и ферментации в дрожжах — алкогольной или молочнокислой ферментации.
При наличии кислорода этот процесс называется аэробным гликолизом — конечным продуктом выступают CO2 и H2O,
Гликолиз представляет собой серию реакций, в которых одна молекула глюкозы распадается на две молекулы пирувата. Но так происходит не всегда, в случаях когда доступ кислорода в этой реакции затруднен, проще говоря, в анаэробных условиях, то пируват превращается в лактат или спирт.
Механизмы анаэробного метаболизма: гликолиз в анаэробных условиях
В анаэробных условиях O2 отсутствует. Отсутствие конечного субстрата O2 ингибирует активность дыхательных структур (до этого они неактивны), и таким образом конечным продуктом распада глюкозы в анаэробных условиях является лактат,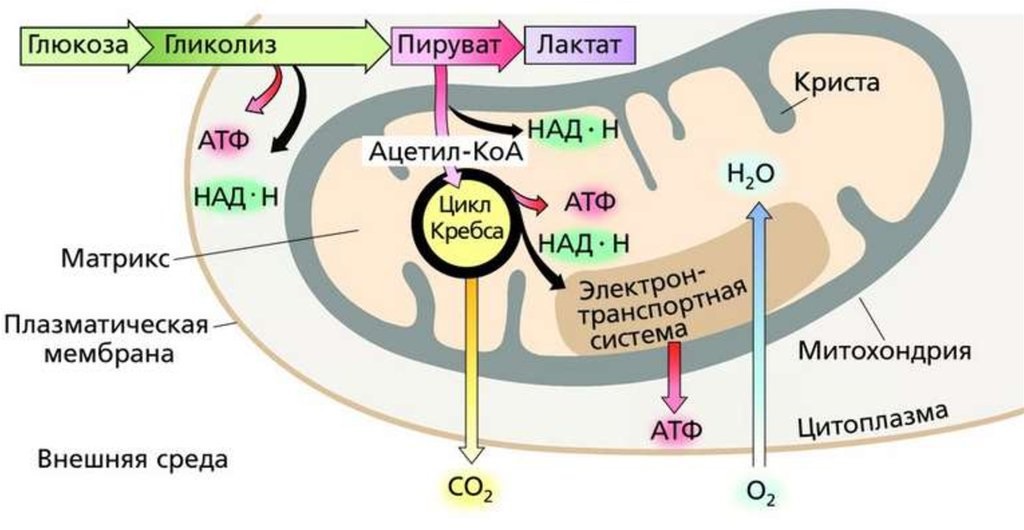
Анаэробный распад глюкозы в физиологических условиях наблюдается в эритроцитах (без митохондрий) и в сильно сокращающихся мышцах (недостаточное кровоснабжение кислородом), это наблюдается при сильной физической работе — присутствует истощение O2 в мышечных клетках (в форме оксимиоглобина).
Лактат в мышечных клетках приводит к мышечной лихорадке, которая преодолевается неинтенсивной мышечной активностью, направленной на полное расщепление лактата в мышцах, до CO2 , H2O и H2 в результате аэробного гликолиза.
При патологических состояниях и онкологических заболеваниях, у многих людей наблюдается так называемый эффект Пастера, при котором клетки из-за уменьшенного поступления крови или кислорода переключаются на анаэробный распад глюкозы, связанный с повышенным ее потреблением (повышенная экспрессия трансмиттеров глюкозы GLUT1 ) и гипогликемией.
Анаэробный гликолиз, как при физиологических, так и при патологических состояниях, характеризуется повышенным уровнем конечного продукта в результате расщепления его лактата в плазме крови — лактатемией, что может привести к ацидозу.
Дальнейшее расщепление лактата
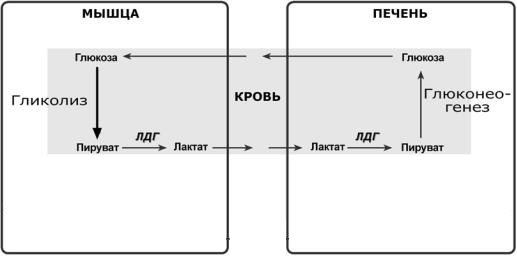
Лактат метаболизируется из печени в пируват, катализируя реакцию в обратном направлении, и глюкоза синтезируется пируватом с использованием ферментов глюконеогенеза в печени через цикл Кори (лактатный цикл).
Вот в этой схеме и кроется ответ на вопрос о том, как влияет анаэробный гликолиз на производство энергии, ведь мы видим, что наша молочная кислота, образовавшаяся в мышцах, через кровь попадает в печень и участвует в процессе глюконеогенеза — создания глюкозы из неуглеводных продуктов.






