Окисление ацетата дает много энергии
Содержание:
Клеточное дыхание
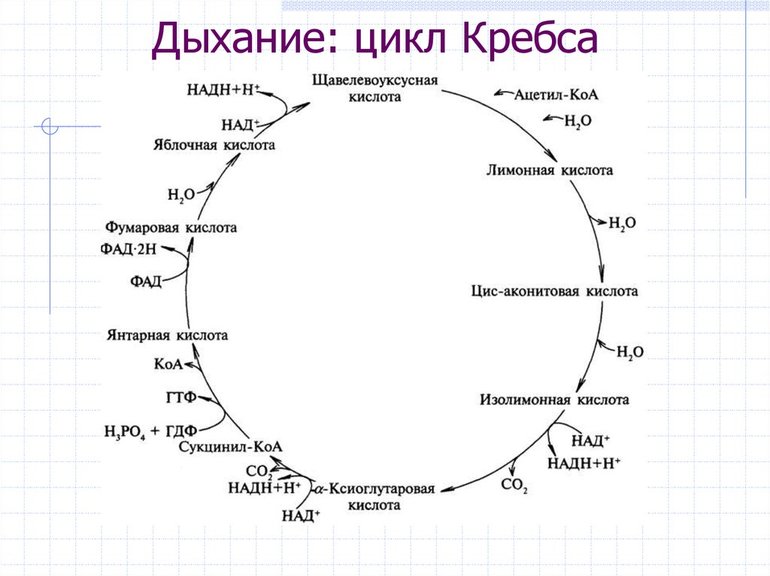
Для нормальной жизнедеятельности живым клеткам постоянно требуется энергия. Ее главный универсальный источник — аденозинтрифосфат (АТФ), способный встраиваться в белки организма напрямую. Это соединение получается в результате ряда реакций окисления, носящих общее название «клеточное дыхание». При этом происходит постепенный распад органических веществ вплоть до простейших неорганических — углекислого газа CO2 и воды H2O.
Структурное строение молекул АТФ содержит фосфорангидридные связи, которые имеют свойство накапливать высвобожденную при прохождении реакций клеточного дыхания энергию, поэтому называются макроэргическими. Так создаются энергетические запасы клеток, которые могут высвобождается при необходимости разрывом этих связей. Процесс синтеза АТФ и класса вспомогательных соединений включает три этапа:
- Гликолиз происходит в цитоплазме.
- В матриксе митохондрий проходят все химические реакции цикла Кребса.
- Окислительное фосфорилирование на внутренней мембране митохондрий.
Преобразование аденозиндифосфата (АДФ) в АТФ характерно для всех этапов. Но наибольшее суммарное количество молекул с макроэргическими связями образуется при фосфорилировании. Это не значит, что процессы гликолиза и ЦК менее важны. Многие соединения, образующиеся во время их протекания, участвуют в регуляции клеточного дыхания.
Регуляция общего пути катаболизма
Осталось поговорить про регуляцию работы пируватдегидрогеназного комплекса и ЦТК. Основная задача для регуляции — поддерживать энергетическое соотношение в клетке. Если в клетке достаточно АТФ, то общий путь катаболизма должен быть отключён. Если концентрация АТФ падает, а нарастает АДФ или АМФ, то нужно запускать общий путь катаболизма.
Основной механизм регуляции пируватдегидрогеназного комплекса и ЦТК — . Она достаточно быстро и чутко реагирует на изменения энергетического соотношения в клетке. Удивительно, что гормональной регуляции с помощью фосфорилирования/дефосфорилирования здесь нет.
Организм отдал регуляцию общего пути катаболизма на усмотрение клетки. Гликолиз и такого доверия не заслужили. С другой стороны, зачем регулировать каждый этап катаболизма гормонально? Достаточно установить контроль над специфическими путями катаболизма, но я отвлёкся.
Регуляция окислительного декарбоксилирования пирувата
Пируватдегидрогеназный комплекс состоит не только из трёх ферментов и пяти коферментов. С первым ферментом (пируватдегидрогеназой) связаны ещё две белковые субъединицы: киназа и фосфатаза. Киназа фосфорилирует пируватдегидрогеназу по -OH группе — переводит её в неактивную форму. Такое случается при избытке АТФ внутри митохондрии. Отдыхаем после тяжёлого дня, лёжа на диване. Фосфатаза же увеличивает свою работоспособность при снижении концентрации АТФ внутри клетки. Это приводит к дефосфорилированию пируватдегидрогеназы — активация фермента. Бежим за маршруткой — нужна энергия.
Киназа и фосфатаза пируватдегидрогеназного комплекса
Это фофорилирование/дефосфорилирование, но не гормональное. Оно подкрепляет аллостерическую регуляцию. Киназа запускается при избытке АТФ, а фосфатаза при его дефиците. Но давайте перейдём к аллостерической регуляции.
Пируватдегидрогеназный комплекс аллостерически ингибируется: АТФ, НАДH, жирными кислотами, ацетил-коэнзимом А.
ПДК ингибируется: АТФ, НАДH, жирными кислотами и ацетил-коэнзимом А
Все эти продукты накапливаются, когда клетке не нужна энергия. Понятно дело, что нужно остановить общий путь катаболизма, если в клетке куча АТФ и субстратов для её синтеза.
Пируватдегидрогеназный комплекс аллостерически активируется: АДФ, НАД+, коэнзимом А, кальцием.
ПДК активируется: АДФ, НАД+, коэнзимом А и кальцием
С первыми тремя всё ясно — они накапливаются, когда клетке нужна энергия, а её не хватает. Нужно ускорить работу пируватдегидрогеназного комплекса — образовать больше ацетил-коэнзима А. Чем больше ацетил-коэнзима А, тем больше энергии будет получено в ходе ЦТК. А что с кальцием? Кальций работает в мышцах и сердце. При сокращениях небольшая часть кальция попадает в митохондрию. Это стимулирует активность пируватдегидрогеназного комплекса и других ферментов ЦТК — об этом ниже.
Регуляция цикла Кребса
Идёт по трём необратимым реакциям: первой, третьей и четвёртой. Регулируются два фермента и один комплекс: цитратсинтаза, изоцитратдегидрогеназа и альфа-кетоглутаратдегидрогеназный комплекс.
Цитратсинтаза. Аллостерически ингибируется: АТФ, НАДH, цитратом и сукцинил-коэнзимом А. Активируется: АДФ.
Изоцитратдегидрогеназа. Ингибируется: АТФ. Активируется: АДФ и кальцием.
Альфа-кетоглутаратдегидрогеназный комплекс. Ингибируется: НАДH и сукцинил-коэнзим А. Активируется: кальцием.
Регуляция ЦТК по первой, третьей и четвертой реакции.
Видно, что наибольшее количество аллостерических регуляторов у цитратсинтазы и пируватдегидрогеназного комплекса. Это неудивительно, две эти реакции отвечают за старт цикла. Цитратсинтаза ингибируется не только продуктом своей реакции, но и продуктом четвертой реакции — сукцинил-коэнзимом А.
Хочешь задать вопрос, похвалить или наговорить гадостей? Тогда залетай в телегу. Там ты сможешь предложить новый формат или разбор темы. А если серьёзно, то эти статьи пишутся для вас, поэтому мне важна обратная связь.
Значение и функции
Этот восьмиэтапный циклический процесс, итогом которого является окисление ацетильного остатка до углекислого газа, может показаться излишне сложным. Тем не менее, он имеет огромное значение в метаболизме промежуточных реакций и выполняет ряд функций. К ним относятся:
- энергетическая;
- анаболическая;
- катаболическая;
- транспортная.
Цикл Кребса участвуют в катаболизме жиров и углеводов. Соединения, образующиеся на разных стадиях процесса, участвуют в синтезе многих необходимых для организма веществ — глутамина, порфиринов, глицина, фенилаланина, цистеина и других. Когда промежуточные продукты покидают ЦК для участия в синтезе, происходит их замещение с помощью так называемых анаплеротических реакций, которые катализируются регуляторными ферментами, например, пируваткарбоксилазой.
Транспортная функция ЦК заключается в содействии гликолизу. Глюкозу невозможно превратить сразу в АТФ, поэтому механизм гликолиза действует поэтапно и сопровождается постоянным перемещением атомов и катионов водорода от одних соединений к другим. Для их транспортировки нужны специальные соединения, которые получаются на одной из стадий ЦТК. Участвующие в гликолизе коферменты цикла Кребса:
- НАД*H+(Никотинамидадениндинуклеотид с катионом водорода). Образуется на III стадии ЦК.
- ФАД*H2 (Флавинадениндинуклеотид с молекулой водорода). Появляется на V стадии ЦК.
Реакции ЦК имеют и большое клиническое значение. Хотя для людей не свойственны мутации, связанные с генами ферментов, участвующих в цикле, однако их редкие проявления губительны для здоровья. Они могут приводить к опухолям мышц и почек, нарушениям работы нервной системы.
Метаболизм
Одно из важнейших свойств живого (вспоминаем биологию) — это обмен веществ с окружающей средой. Действительно, только живое существо может что-то поглощать из окружающей среды, и что-то потом в неё выделять.
В биохимии обмен веществ принято называть «метаболизм». Обмен веществ, обмен энергией с окружающей средой — это метаболизм.
Когда мы, допустим, съели бутерброд с курицей, мы получили белки (курица) и углеводы (хлеб). В процессе пищеварения белки распадутся до аминокислот, а углеводы — до моносахаров. То, что я описал сейчас, называется катаболизм, то есть распад сложных веществ на более простые. Первая часть метаболизма — это катаболизм.
Ещё один пример. Ткани в нашем организме постоянно обновляются. Когда отмирает старая ткань, её обломки растаскивают макрофаги, и они заменяется новой тканью. Новая ткань создаётся в процессе синтеза белка из аминокислот. Синтез белка происходит в рибосомах. Создание нового белка (сложного вещества) из аминокислот (простого вещества) — это анаболизм.
Итак, анаболизм — это противоположность катаболизму. Катаболизм — это разрушение веществ, анаболизм — это создание веществ. Кстати, чтобы их не путать, запомните ассоциацию: «Анаболики. Кровью и потом». Это голливудский фильм (довольно скучный, на мой взгляд) о спортсменах, применяющих анаболики для роста мышц. Анаболики — рост, синтез. Катаболизм — обратный процесс.
Примечания и ссылки
(in) ,
1937 г.(доступ 7 ноября 2015 г. ) : « Нобелевская премия по физиологии и медицине 1937 г. была присуждена Альберту Сент-Дьёрдьи» за его открытия, связанные с процессами биологического горения, с особым упором на витамин С и катализ фумаровой кислоты ». » .
(in) (по состоянию на 7 ноября 2015 г. )Нобелевская премия по физиологии и медицине 1953 г. была разделена поровну между Гансом Адольфом Кребсом» за открытие им лимонной кислоты
кислотный цикл »и Фриц Альберт Липманн« за открытие кофермента А и его важность для промежуточного метаболизма » .
(in) Х. Гест , Эволюционные корни цикла лимонной кислоты у прокариот , Симпозиум Биохимического общества , вып
54,
1987 г., стр. 3–16 ( PMID )
(in) Энрике Мелендес-Хевиа, Томас Г. Уодделл и Марта Касканте , Загадка цикла лимонной кислоты Кребса: сборка частей химически осуществимых реакций и оппортунизм в разработке метаболических путей во время эволюции , Journal of Molecular Эволюция , т. 43, п о 1,
Сентябрь 1996, стр. 293-303
(in) Оливер Эбенхох и Рейнхарт Генрих , Эволюционная оптимизация метаболических путей. Теоретическая реконструкция стехиометрии систем, продуцирующих АТФ и НАДН , Бюллетень математической биологии , вып. 63, п о 1,
Январь 2001 г., стр. 21-55
↑ и (в) Takehiko САХАРЕ, Ясухиро~d Такаде, Ёдзи Takeuchi Наото Ямаокие и Нориюки Fukunaga , Клонирование, секвенирование и экспрессии гена , кодирующий мономерных изоцитратдегидрогеназу из азотфиксирующей Бактерии, Азотобактер ьтеШого , Bioscience, Biotechnology , и Биохимия , т. 66, п о 3,
Март 2002 г., стр. 489-500
↑ и (ru) Мишель Э. ван дер Рест, Кристиан Франк и Доув Моленаар , Функции мембранно-ассоциированных и цитоплазматических малатдегидрогеназ в лимоннокислотном цикле Escherichia coli , Journal of bacteriology , vol. 182, п о 24,
декабрь 2000, стр. 6892-6899
(in) Р.К. Портер и М.Д. Бранд , Проводимость митохондриальных протонов и отношения H + / O не зависят от скорости транспорта электронов в изолированных гепатоцитах , Biochemical Journal , vol. 310,
1995 г., стр. 379–382
(in) Дэвид О. Ламбет, Кристин Н. Тьюс, Стивен Адкинс, Дин Фролих и Барри И. Милавец , Экспрессия двух сукцинил-КоА-синтетаз с нуклеотидами разной специфичности в тканях млекопитающих , Журнал биологической химии , вып. 279, п о 35,
27 августа 2004 г., стр. 36621-36624
(in) Элвуд А. Маллинс, Джули А. Франсуа и Джозеф Т. Каппок , Специализированный цикл лимонной кислоты, требующий сукцинил-кофермента А (КоА): трансфераза ацетат-КоА (AARC) придает устойчивость к уксусной кислоте ацидофильному грибку Acetobacter aceti , Журнал бактериологии , вып. 190, п о 14,
июль 2008 г., стр. 4933-4940
(in) Ирен Э. Кортези-Теулаз Габриэла Э. Бергонцелли, Генри Хьюз, Даниэль Бахманн Даниэль Ф. Шордерет Андре Л. Блюм и Николас Л. Орнстон , Клонирование и характеристика сукцинил-КоА Helicobacter pylori : трансфераза ацетоацетат-КоА, роман Прокариотический член семейства CoA-трансфераз , Journal of Biological Chemistry , vol. 272, п о 41,
10 октября 1997 г., стр. 25659-25667
(в) Энтони Д. Baughn, Скотт J. Garforth, Vilchèze Кэтрин и William R. Jacobs Jr. , анаэробного типа α-кето ферредоксина оксидоредуктазы завершает Окислительный трикарбоновых кислот цикл микобактерий туберкулеза , PLoS патогенам , вып. 5, п о 11,
ноябрь 2009 г., e1000662
(in) Шуйи Чжан и Дональд А. Брайант , Цикл трикарбоновых кислот у цианобактерий , Science , vol. 334, п о 6062,
16 декабря 2011 г., стр. 1551-1553
(in) Максим В. Иванников и Грегори Т. Маклауд , Уровни свободного Ca2 + в митохондриях и их влияние на энергетический метаболизм в терминалах моторных нервов дрозофилы , Biophysical Journal , Vol. 104, п о 11,
4 июня 2013 г., стр. 2353-2361
(in) Ричард Дентон, Филип Дж. Рэндл, Барбара Дж. Бриджес, Рональд Х. Купер, Алан Л. Керби, Хелен Т. Паск, Дэвид Л. Северсон, Дэвид и Сьюзан Уайтхаус Стэнсби , Регулирование пируватдегидрогеназы млекопитающих , Молекулярная и клеточная биохимия , т. 9, п о 1,
Октябрь 1975 г., стр. 27-53
(ru) Ингибирование гидроксилаз индуцируемого гипоксией фактора (HIF) промежуточными продуктами цикла лимонной кислоты. ВОЗМОЖНЫЕ СВЯЗИ МЕЖДУ КЛЕТОЧНЫМ МЕТАБОЛИЗМОМ И СТАБИЛИЗАЦИЕЙ HIF , Пеппи Койвунен, Майя Хирсиля, Анне М. Ремес, Ильмо Э. Хассинен, Кари И. Кивирикко и Йоханна Мюллюхарью , журнал биологической химии , вып. 282, п о 7,
16 февраля 2007 г., стр. 4524-4532
(in) Премжит П. Халарнкар и Гэри Дж. Бломквист , Сравнительные аспекты метаболизма пропионата , Сравнительная биохимия и физиология, Часть B: Сравнительная биохимия , том. 92, п о 2
1989 г., стр. 227-231
Пятая реакция
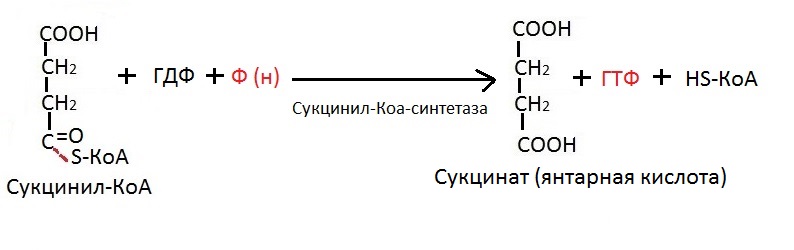
В прошлом шаге мы говорили, что сукцинил-КоА — это эфир янтарной кислоты. А теперь мы получим саму янтарную кислоту, то есть сукцинат, из сукцинила-КоА. Крайне важный момент: именно в этой реакции происходит субстратное фосфорилирование.
Фосфорилирование вообще (оно бывает окислительное и субстратное) — это добавление фосфорной группы PO3 к ГДФ или АТФ, чтобы получить полноценный ГТФ, или соответственно, АТФ. Субстратное отличается тем, что эта самая фосфорная группа отрывается от какого-либо вещества, её содержащую. Ну проще говоря, она переносится с СУБСТРАТА на ГДФ или АДФ. Поэтому и называется — «субстратное фосфорилирование».
Ещё раз: на момент начала субстратного фосфорилирования у нас имеется дифосфатная молекула — гуанозинДифосфат или аденозинДифосфат. Фосфорилирование заключается в том, что молекула с двумя остатками фосфорной кислоты — ГДФ или АДФ «достраивается» до молекулы с тремя остатками фосфорной кислоты, чтобы получились гуанозинТРИфосфат или аденозинТРИфосфат. Этот процесс происходит во время превращения сукцинила-КоА в сукцинат (то есть, в янтарную кислоту).
На схеме вы можете увидеть буквы Ф (н). Это значит «неорганический фосфат». Неорганический фосфат переходит от субстрата на ГДФ, чтобы в продуктах реакции был хороший, полноценный ГТФ. Теперь давайте посмотрим на саму реакцию:
Интеграция с основными метаболическими путями
Анаплеротические и катаплеротические маршруты
Несколько метаболических путей сходятся в цикле Кребса. Большинство этих реакций производят метаболиты, участвующие в цикле, и поэтому являются анаплеротическими реакциями ; процессы, которые, с другой стороны, потребляют метаболиты цикла Кребса, считаются катаплеротическими.
Все промежуточные продукты цикла Кребса, такие как цитрат , изоцитрат , α-кетоглутарат , сукцинат , фумарат , L- малат и оксалоацетат , регенерируются на каждом этапе цикла. Увеличение доступного количества одного из этих метаболитов увеличивает общее количество всех метаболитов в цикле, поскольку каждый промежуточный продукт постепенно превращается во все другие промежуточные продукты в цикле. Это причина того, почему производство любого из метаболитов цикла Кребса имеет общий анаплеротический эффект в этом цикле, в то время как потребление любого одного метаболита имеет катаплеротический эффект в целом цикла.
Молекулы пирувата , образующиеся в результате гликолиза , активно транспортируются из цитозоля в матрикс митохондрий через внутреннюю мембрану митохондрий . Попав в матрицу, они могут окисляться и реагировать с коферментом А с образованием CO 2., Ацетил-КоА и НАДН , или они могут быть карбоксилирования с помощью пируваткарбоксилазы с образованием оксалоацетата . Это анаплеротическая реакция, которая увеличивает поток и, следовательно, поток через цикл Кребса, когда клетка сталкивается с повышенной потребностью в метаболической энергии, например, в миоцитах .
Циклическое кормление с ацетил-КоА
Ацетил-КоА , однако, происходит от окисления из пирувата или из бета-окисления из жирных кислот , никогда не регенерируют цикла Кребса: наоборот, каждый виток из потребляет цикла одна молекула ацетил-КоА в оксалоацетат молекулы митохондриального матрикса, в то время как ацетильный остаток ацетил-КоА полностью окисляется до CO и в H 2 Oчерез дыхательную цепь , позволяя восстанавливать метаболическую энергию в форме АТФ путем окислительного фосфорилирования .
Следовательно, метаболические пути, которые производят или потребляют ацетил-КоА, не являются анаплеротическими или катаплеротическими для цикла Кребса.
Глюкогенез
В печени карбоксилирование цитозольного пирувата до митохондриального оксалоацетата является продвинутым этапом глюконеогенеза , который превращает лактат и дезаминированный аланин в глюкозу под действием повышенного уровня глюкагона и / или адреналина в крови . В этих условиях образование оксалоацетата в митохондриях не проявляет анаплеротического эффекта, поскольку митохондриальный L -малат расходуется с образованием цитозольного оксалоацетата, который превращается в глюкозу.
Деградация белков
В процессе деградации из белков , что полипептидные цепи являются расщепляются с помощью пептидаз в аминокислотах , которые составляют их. Тогда их углеродная цепь может:
- войти в цикл Кребса — как, например , в случае глутамата и глутамина , которые входят в виде -кетоглутарата — который представляет собой заживляющий эффект , — мы говорим о glucoforming аминокислоты , потому что они могут быть ориентированы на глюконеогенез с помощью в L -malate ;
- превращаться в ацетил-КоА для окисления и производства энергии, как в случае с лейцином , изолейцином , лизином , фенилаланином , триптофаном и тирозином ;
- превращаться в кетоновые тела , способные либо окисляться тканями, отличными от печени, в которой они производятся, либо выводиться с мочой или дыханием — это называется кетоформованными аминокислотами .
Деградация липидов
В процессе липолиза , триглицериды которые гидролизуют в глицерина и жирных кислот . В печени , глицерин может быть преобразован в глюкозу с помощью в фосфатном диоксиацетон и глицеральдегид — 3-фосфата в контексте глюконеогенеза . Во многих тканях , особенно в сердце и скелетных мышцах , жирные кислоты разлагаются посредством β-окисления , в результате чего образуется митохондриальный ацетил-КоА, способный присоединиться к циклу Кребса. Жирные кислоты с нечетным числом атомов от углерода продукция пропионила-КоА , который превращается в сукциниле-СоА и присоединяется цикл Кребса , как промежуточный заживляющие.
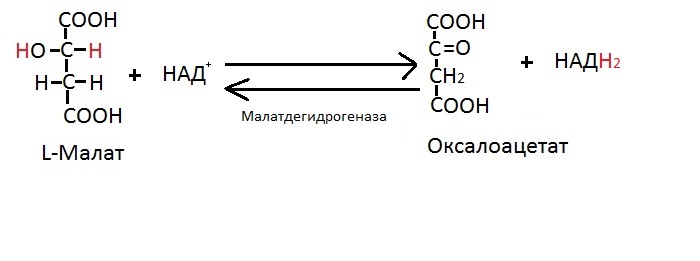
Восьмая реакция
Последняя реакция Цикла Кребса — это окисление L-яблочной кислоты до оксалоацетата, то есть до щавелевоуксусной кислоты. Как вы поняли, «оксалоацетат» и «щавелевоуксусная кислота» — это синонимы. Вы, наверное, помните, что щавелевоуксусная кислота является компонентом первой реакции цикла Кребса.
Здесь же отмечаем особенность реакции: образование НАДH2, который понесёт электроны в дыхательную цепь. Не забудьте также реакции 3,4 и 6, там также образуются переносчики электронов и протонов для дыхательной цепи.
Как видите, я специально выделил красным цветом реакции, в ходе которых образуются НАДH и ФАДH2. Это очень важные вещества для дыхательной цепи. Зелёным я выделил реакцию, в рамках которой происходит субстратное фосфорилирование, и получается ГТФ.
Точка пересечения распада и синтеза.
Цикл Кребса как ступень катаболизма.
Как связаны метаболизм и цикл Кребса? Дело в том, что именно цикл Кребса является одной из важнейших точек, в которой сходятся пути анаболизма и катаболизма. Именно в этом и заключается его значение.
Давайте разберём это на схемках. Катаболизм можно условно представить как расщепление белков, жиров и углеводов в нашей пищеварительной системе. Итак, мы скушали пищу из белков, жиров, и углеводов, что дальше?
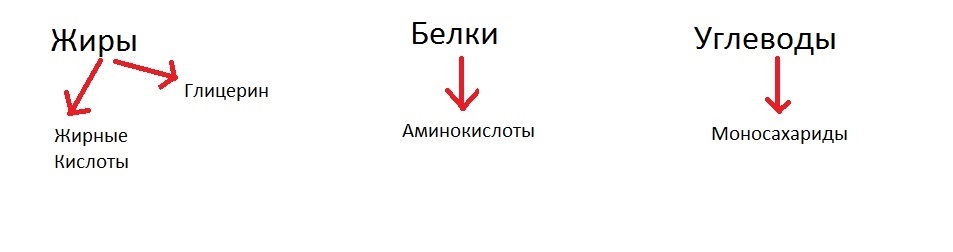
А дальше все эти вещества распадутся на простые составляющие:
- Жиры — на глицерин и жирные кислоты (могут быть и другие компоненты, я решил взять самый простой пример);
- Белки — на аминокислоты;
- Полисахаридные молекулы углеводов — на одинокие моносахариды.
Далее, в цитоплазме клетки, последует превращение этих простых веществ в пировиноградную кислоту (она же — пируват). Из цитоплазмы пировиноградная кислота попадёт в митохондрию, где превратится в ацетил коэнзим А. Пожалуйста, запомните эти два вещества — пируват и ацетил КоА, они очень важны.
Давайте теперь посмотрим, как происходит этап, который мы сейчас расписали:
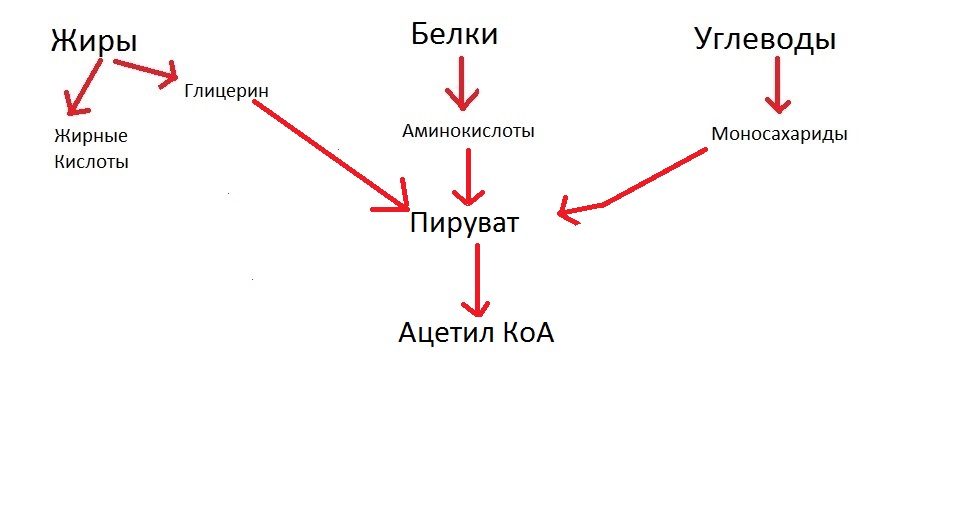
Важная деталь: аминокислоты могут превращаться в ацетил КоА сразу, минуя стадию пировиноградной кислоты. Жирные кислоты сразу превращаются в ацетил КоА. Учтём это и подредактируем нашу схемку, чтобы получилось правильно:
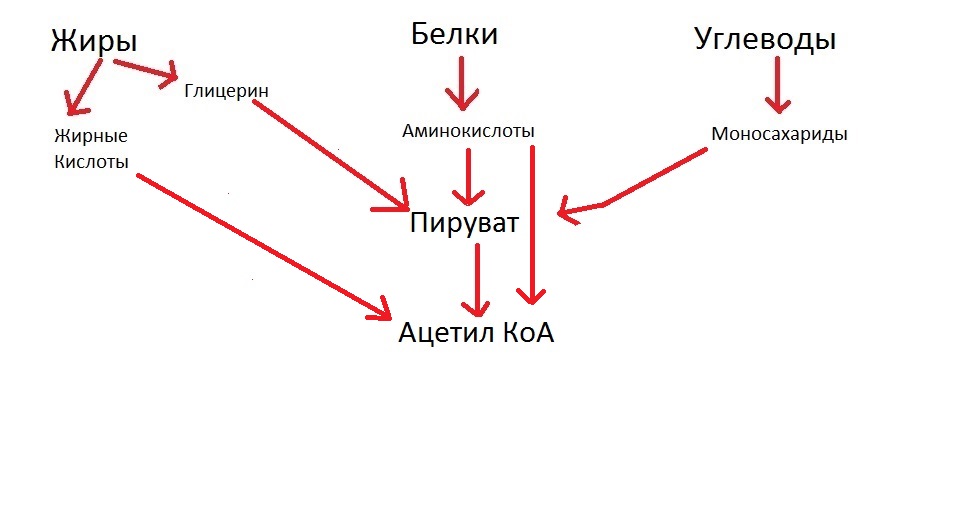
Превращения простых веществ в пируват происходят в цитоплазме клеток. После этого пируват поступает в митохондрии, где успешно превращается в ацетил КоА.
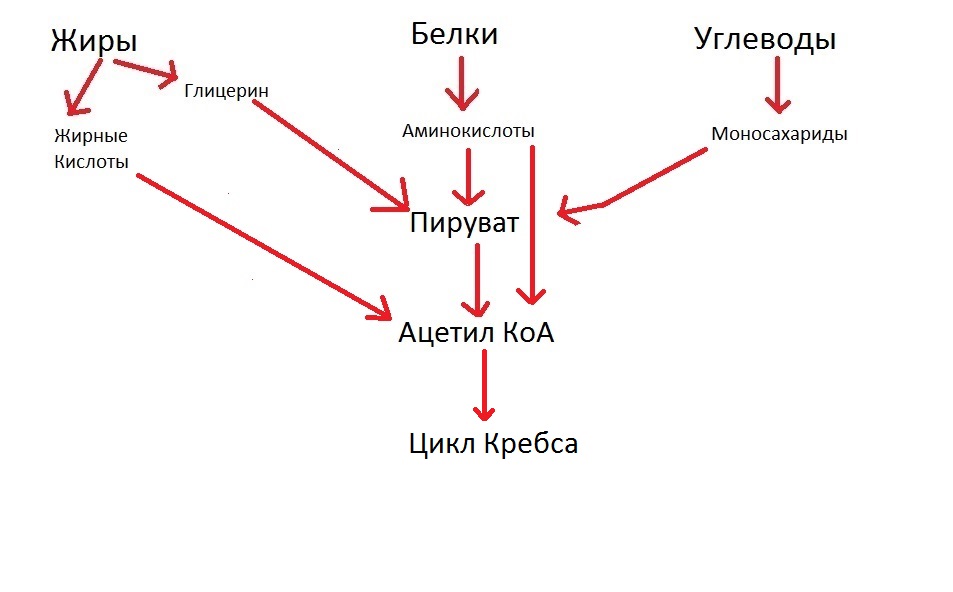
Для чего пируват превращается в ацетил КоА? Именно для того, чтобы запустить наш цикл Кребса. Таким образом, мы можем сделать ещё одну надпись в схеме, и получится правильная последовательность:
В результате реакций цикла Кребса образуются важные для жизнедеятельности вещества, главные из которых:
- НАДH (НикотинАмидАденинДиНуклеотид+ катион водорода) и ФАДH2 (ФлавинАденинДиНуклеотид+молекула водорода). Я специально выделил заглавными буквами составные части терминов, чтобы легче было читать, в норме их пишут одним словом. НАДH и ФАДH2 выделяются в ходе цикла Кребса, чтобы потом принять участие в переносе электронов в дыхательную цепь клетки. Иными словами, эти два вещества играют важнейшую роль в клеточном дыхании.
- АТФ, то есть аденозинтрифосфат. Это вещество имеет две связи, разрыв которых даёт большое количество энергии. Этой энергией снабжаются многие жизненно важные реакции;
Также выделяются вода и углекислый газ. Давайте отразим это на нашей схеме:
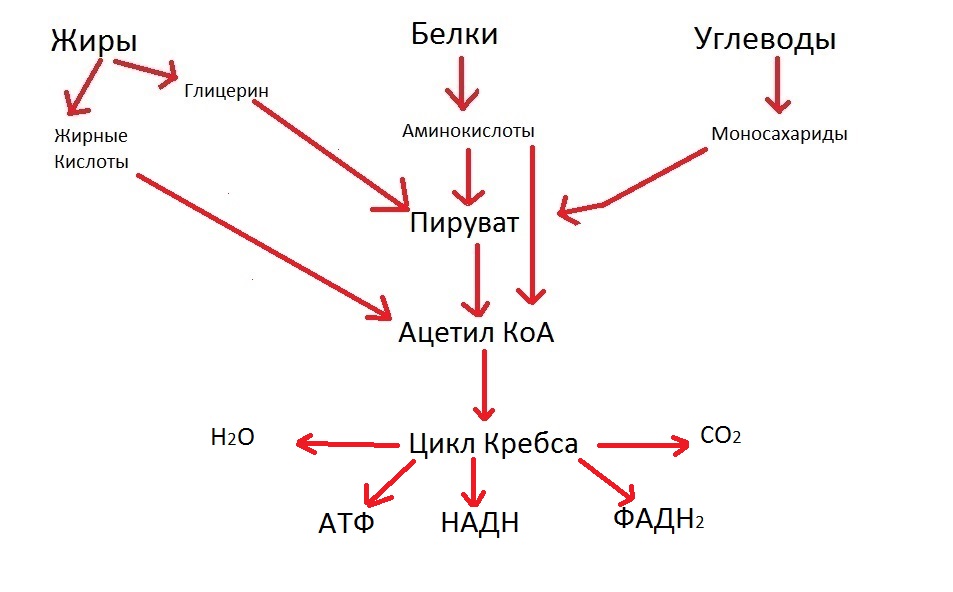
Кстати, весь цикл Кребса происходит в митохондриях. Именно там, где проходит и подготовительный этап, то есть превращение пирувата в ацетил КоА. Не зря кстати митохондрии называют «энергетическая станция клетки».
Цикл Кребса как начало синтеза
Цикл Кребса удивителен тем, что он не только даёт нам ценные АТФ (энергию) и коферменты для клеточного дыхания. Если посмотрите на предыдущую схему, вы поймёте, что цикл Кребса — это продолжение процессов катаболизма. Но вместе с тем он является и первой ступенькой анаболизма. Как это возможно? Как один и тот же цикл может и разрушать, и создавать?
Оказывается, отдельные продукты реакций цикла Кребса могут частично отправляться на синтез новых сложных веществ в зависимости от потребностей организма. Например, на глюконеогенез — это синтез глюкозы из простых веществ, не являющихся углеводами.
Ещё раз:
- Реакции цикла Кребса каскадны. Они происходят одна за другой, и каждая предыдущая реакция запускает последующую;
- Продукты реакций цикла Кребса частично идут на запуск последующей реакции, а частично — на синтез новых сложных веществ.
Давайте попробуем отразить это на схеме, чтобы цикл Кребса был обозначен именно как точка пересечения распада и синтеза.

Голубыми стрелочками я отметил пути анаболизма, то есть создания новых веществ. Как видите, цикл Кребса действительно является точкой пересечения многих процессов и разрушения, и созидания.







