Б. аэробный гликолиз
Содержание:
- Содержание:
- История исследования
- Обходной путь 3. Образование глюкозы
- ЛокализацияПравить
- Этапы окислительного гликолиза. Фаза 1.
- Бескислородное окисление глюкозы включает два этапа
- Анаэробный гликолиз глюкозы в эритроцитах, 2,3-БФГ и эффект БораПравить
- Общая характеристика систем энергообеспечения мышечной деятельности.
- Бескислородный этап энергетического обмена
- Окисление
- Что такое гликолиз
- Энергетический обмен
- Стадия реакции окисления глюкозы
- Теги
- Значение гликолиза
Содержание:
Аэробный против анаэробного гликолиза
Гликолиз — это первая стадия образования АТФ, которая происходит в цитозоле вне митохондрий, с использованием глюкозы в качестве источника энергии. Это происходит как в аэробной, так и в анаэробной среде и является единственным путем, способным производить АТФ в отсутствие кислорода. Следовательно, его можно увидеть в организмах, таких как прокариоты, клетках, таких как эритроциты, и в гипоксической среде, такой как быстро сокращающаяся мышечная ткань или ишемическая ткань, в которой отсутствуют митохондрии. Процесс гликолиза можно разделить на аэробный или анаэробный гликолиз, в зависимости от наличия кислорода в окружающей среде. Однако в обоих процессах исходным источником является глюкоза, а конечным продуктом — пируват.

(Источник изображения: «Анаэробные и аэробные пути» SparkNotes.com. SparkNotes LLC. N.d .. Web. 13 сентября 2013 г.)
Аэробный гликолиз
Аэробный гликолиз — это гликолитический путь, который происходит в цитозоле в присутствии кислорода. По сравнению с анаэробным гликолизом этот путь намного более эффективен и производит больше АТФ на молекулу глюкозы. При аэробном гликолизе конечный продукт, пируват, переносится в митохондрии для запуска цикла лимонной кислоты. Следовательно, конечными продуктами аэробного гликолиза являются 34 молекулы АТФ, вода и углекислый газ.
Анаэробный гликолиз
Анаэробный гликолиз происходит в цитоплазме, когда клетке не хватает оксигенированной среды или митохондрий. В этом случае НАДН окисляется в цитозоле до НАД +, превращая пируват в лактат. Анаэробный гликолиз производит (2 лактата + 2 АТФ + 2 H2O + 2 H +) из одной молекулы глюкозы. В отличие от аэробного гликолиза, анаэробный гликолиз производит лактат, который снижает pH и инактивирует ферменты.
В чем разница между аэробным и анаэробным гликолизом?
• Аэробный гликолиз происходит в среде, богатой кислородом, тогда как анаэробный гликолиз происходит в среде с недостатком кислорода.
• Аэробный гликолиз более эффективен, чем анаэробный гликолиз; следовательно, он производит большее количество АТФ, чем анаэробный гликолиз.
• Аэробный гликолиз происходит только у эукариот, в то время как анаэробный гликолиз происходит как у прокариот, так и у эукариот.
• В отличие от анаэробного гликолиза, конечный продукт аэробного гликолиза (пируват) используется для инициирования других путей в митохондриях.
• Анаэробный гликолиз производит 2 АТФ на молекулу глюкозы, в то время как аэробный гликолиз производит от 36 до 38 АТФ на молекулу глюкозы.
• Конечным конечным продуктом анаэробного гликолиза является лактат, который может быть вредным для самой клетки, тогда как продукт аэробного гликолиза — это вода и углекислый газ, которые не вредны для клеток.
• В отличие от анаэробного гликолиза, НАДН + Н + подвергаются окислительному фосфорилированию в присутствии кислорода при аэробном гликолизе.
• Пируват восстанавливается до лактата во время анаэробного гликолиза, тогда как во время аэробного гликолиза пируват окисляется до ацетилкофермента А (ацетил-КоА).
История исследования
Гликолиз первый из открытых и наиболее исследован метаболический путь. 1897 братья Ганс и Эдуард Бюхнер занимались изготовлением бесклеточной экстрактов дрожжей для терапевтического применения. Очевидно, они не могли использовать ядовитые для человека антисептики, как фенол, поэтому попытались распространен в кулинарии консервант — сахарозу. Выяснилось, что в дрожжевых экстрактах это вещество быстро зброджувалась к этиловому спирту. Так впервые установлено, что брожение может происходить вне живых клеток. В 1907 году Эдуарду Бюхнеру была присуждена Нобелевская премия по химии.
Со времени открытия внеклеточного брожения до 1940-х годов исследования реакций гликолиза было одной из основных задач биохимии. Описанием этого метаболического пути в клетках дрожжей занимались Отто Варбург, Ганс фон Эйлер-Хельпин и Артур Гарден (два последних получили Нобелевскую премию по химии 1929 года), в мышцах — Густав Эмбден и Отто Меергоф (Нобелевская премия по медицине и физиологии 1922 ). Также свой вклад в исследование гликолиза сделали Карл Нойберг, Яков Парнас, Герти и Карл Кори.
Важными «побочными» открытиями, сделанными благодаря изучению гликолиза, была разработка многих методов очистки ферментов, выяснения центральной роли АТФ и других фосфорильованих соединений в метаболизме, открытие коэнзимов, таких как НАД.
Обходной путь 3. Образование глюкозы
От глюкозо-6-фосфата отщепляется фосфатная группа. Катализирует реакцию глюкозо-6-фосфатаза.
Образование глюкозы в третьем обходном пути
Но этот фермент не может находиться в цитоплазме. Почему? Тогда клетка не смогла бы выполнять свои функции. Представьте, что глюкоза попадает в печень. Она сразу же фосфорилируется с помощью глюкокиназы. Так клетка добивается двух вещей:
- Увеличивает реакционную способность глюкозы;
- Не даёт глюкозе выбраться из клетки. Потому что для глюкозо-6-фосфата нет белков переносчиков в цитоплазме. А вот для обычной глюкозы есть —
Если мы засунем в эту систему фермент глюкозо-6-фосфатазу, то она будет постоянно превращать глюкозо-6-фосфат в глюкозу. А та будет вылетать из клетки. Получается, что вся система работает в холостую. Поэтому глюкозо-6-фосфатаза не работает в цитоплазме, а находится в эндоплазматическом ретикулуме (ЭПР). Он отделён от цитоплазмы биологической мембраной.
В мембране ЭПР есть белки переносчики для глюкозо-6-фосфата. Он переносится в ЭПР, где дефосфорилируется глюкозо-6-фосфатазой. Образуется глюкоза и неорганический фосфат. Они переносятся в цитоплазму, опять же с помощью своих белков переносчиков. А дальше глюкоза попадает в кровь через глюкозный транспортёр второго типа.
Глюкозо-6-фосфатаза работает в эндоплазматическом ретикулуме. Так клетка избегает холостых реакций.
Вот и весь глюконеогенез. Держите общую картинку.
Пируваткарбоксилаза не влезла, извиняйте.
ЛокализацияПравить
В клетках эукариотических организмов десять ферментов, катализирующих распад глюкозы до ПВК, находятся в цитозоле, все остальные ферменты, имеющие отношение к энергетическому обмену, — в митохондриях. Поступление глюкозы в клетку осуществляется двумя путями: натрий-зависимый ко-транспорт (преимущественно для энтероцитов и эпителия почечных канальцев) и облегчённая диффузия глюкозы с помощью белков-переносчиков. Работа этих белков-транспортёров контролируется гормонами и, в первую очередь, инсулином. Сильнее всего инсулин стимулирует транспорт глюкозы в мышцах и жировой ткани.
Этапы окислительного гликолиза. Фаза 1.
В аэробном гликолизе выделяются 2 фазы.
Первая фаза – подготовительная. В ней глюкоза вступает в реакцию с 2 молекулами АТФ. Эта фаза состоит из 5 последовательных ступеней биохимических реакций.
1-я ступень. Фосфорилирование глюкозы
Фосфорилирование, то есть процесс переноса остатков фосфорной кислоты в первой и последующих реакциях производится за счет молекул адезинтрифосфорной кислоты.
В первой ступени остатки фосфорной кислоты из молекул адезинтрифосфата переносятся в молекулярную структуру глюкозы. В ходе процесса получается глюкозо-6-фосфат. В качестве катализатора в процессе выступает гексокиназа, ускоряющая процесс с помощью ионов магния, выступающих в качестве кофактора. Ионы магния задействованы и в других реакциях гликолиза.

2-я ступень. Образование изомера глюкозо-6-фосфата

На 2-й ступени происходит изомеризация глюкозо-6-фосфата во фруктозу-6-фосфат.
Изомеризация – образование веществ, имеющих одинаковый вес, состав химических элементов, но обладающих разными свойствами вследствие различного расположения атомов в молекуле. Изомеризация веществ осуществляется под действием внешних условий: давления, температур, катализаторов.
В данном случае процесс осуществляется под действием катализатора фосфоглюкозоизомеразы при участии ионов Mg+.
3-я ступень. Фосфорилирование фруктозо-6-фосфата
На данной ступени происходит присоединение фосфорильной группы за счет АТФ. Процесс осуществляется при участии фермента фосфофруктокиназа-1. Этот фермент и предназначен только для участия в гидролизе. В результате реакции получаются фруктозо-1,6-бисфосфат и нуклеотид адезинтрифосфат.

АТФ – адезинтрифосфат, уникальный источник энергии в живом организме. Представляет собой довольно сложную и громоздкую молекулу, состоящую из углеводородных, гидроксильных групп, азота и групп фосфорной кислоты с одной свободной связью, собранных в нескольких циклических и линейных структурах. Высвобождение энергии происходит в результате взаимодействия остатков фосфорной кислоты с водой. Гидролиз АТФ сопровождается образованием фосфорной кислоты и выделением 40-60 Дж энергии, которую организм затрачивает на свою жизнедеятельность.
Но прежде должно произойти фосфорилирование глюкозы за счет молекулы Адезинтрифосфата, то есть перенос остатка фосфорной кислоты в глюкозу.
4-я ступень. Распад фруктозо-1,6-дифосфата

В четвертой реакции фруктозо-1,6-дифосфат распадается на два новых вещества.
- Диоксиацетонфосфат,
- Глицеральд альдегид-3-фосфат.
В данном химическом процессе в качестве катализатора выступает альдолаза, фермент, участвующий в энергетическом обмене, и необходимый при диагностировании ряда заболеваний.
5-я ступень. Образование триозофосфатных изомеров
И, наконец, последний процесс – изомеризация триозофосфатов.

Глицеральд-3-фосфат продолжит участвовать в процессе аэробного гидролиза. А второй компонент – диоксиацетон фосфат при участии фермента триозофосфатизомеразы преобразуется в глицеральдегид-3-фосфат. Но трансформация эта – обратимая.
Бескислородное окисление глюкозы включает два этапа
В анаэробном процессе пировиноградная кислота восстанавливается до молочной кислоты (лактата), поэтому в микробиологии анаэробный гликолиз называют молочнокислым брожением. Лактат является метаболическим тупиком и далее ни во что не превращается, единственная возможность утилизовать лактат – это окислить его обратно в пируват.
Многие клетки организма способны к анаэробному окислению глюкозы. Для эритроцитов он является единственным источником энергии.
Клетки скелетной мускулатуры за счет бескислородного расщепления глюкозы способны выполнять мощную, быструю, интенсивную работу, как, например, бег на короткие дистанции, напряжение в силовых видах спорта.
Вне физических нагрузок бескислородное окисление глюкозы в клетках усиливается при гипоксии – при различного рода анемиях, при нарушении кровообращения в тканях независимо от причины.
Первый этап гликолиза – подготовительный, здесь происходит затрата энергии АТФ, активация глюкозы и образование из нее триозофосфатов.
Первая реакция гликолиза сводится к превращению глюкозы в реакционно-способное соединение за счет фосфорилирования 6-го, не включенного в кольцо, атома углерода. Эта реакция является первой в любом превращении глюкозы, катализируется гексокиназой.
Вторая реакция необходима для выведения еще одного атома углерода из кольца для его последующего фосфорилирования (фермент глюкозофосфат-изомераза). В результате образуется фруктозо-6-фосфат.
Третья реакция – фермент фосфофруктокиназа фосфорилирует фруктозо-6-фосфат с образованием почти симметричной молекулы фруктозо-1,6-дифосфата. Эта реакция является главной в регуляции скорости гликолиза.
В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат-альдолазой с образованием двух фосфорилированных триоз-изомеров – альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ).
Пятая реакция подготовительного этапа – переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга при участии триозофосфатизомеразы. Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97%, доля глицеральдегидфосфата – 3%. Эта реакция, при всей ее простоте, определяет дальнейшую судьбу глюкозы:
- при нехватке энергии в клетке и активации окисления глюкозы диоксиацетонфосфат превращается в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза,
- при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний отправляется на синтез жиров.
Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ.
Шестая реакция гликолиза (фермент глицеральдегидфосфат-дегидрогеназа) – окисление глицеральдегидфосфата и присоединение к нему фосфорной кислоты приводит к образованию макроэргического соединения 1,3-дифосфоглицериновой кислоты и НАДН.
В седьмой реакции (фермент фосфоглицераткиназа) энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тратится на образование АТФ.
Реакция получила дополнительное название – реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (от субстрата реакции) в отличие от окислительного фосфорилирования (от электрохимического градиента ионов водорода на мембране митохондрий).
Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы изомеризуется в 2-фосфоглицерат.
Девятая реакция – фермент енолаза отрывает молекулу воды от 2-фосфоглицериновой кислоты и приводит к образованию макроэргической фосфоэфирной связи в составе фосфоенолпирувата.
Десятая реакция гликолиза – еще одна реакция субстратного фосфорилирования – заключается в переносе пируваткиназой макроэргического фосфата с фосфоенолпирувата на АДФ и образовании пировиноградной кислоты.
Последняя реакция бескислородного окисления глюкозы, одиннадцатая – образование молочной кислоты из пирувата под действием лактатдегидрогеназы
Важно то, что эта реакция осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, образующийся в 6-й реакции, в отсутствие кислорода не может окисляться в митохондриях
| У плода и детей первых месяцев жизни преобладает анаэробный распад глюкозы, в связи с чем концентрация молочной кислоты в крови у них выше чем у взрослых. |
| При наличии кислорода пировиноградная кислота переходит в митохондрию и превращается в ацетил-S-КоА. |
Анаэробный гликолиз глюкозы в эритроцитах, 2,3-БФГ и эффект БораПравить
Рис. 18.1. Анаэробный гликолиз в эритроцитах
Рис. 18.2. Образование 2,3-бисфосфоглицерата в эритроцитах
Эритроциты транспортируют большое количество кислорода, которым снабжают все органы человека. По иронии судьбы сами они не могут использовать этот кислород. Источником энергии для эритроцитов служит глюкоза, при окислении которой в ходе анаэробного гликолиза в эритроцитах образуется АТФ (рис. 18.1). Аэробное окисление глюкозы в эритроцитах невозможно, так как в них отсутствуют митохондрии и вместе с ними все ферменты цикла Кребса. Кроме того, в них отсутствуют также ферменты, необходимые для окисления жирных кислот и использования кетоновых тел.
Функция эритроцитов — транспорт кислорода ко всем органам и тканям организма. Гемоглобин эритроцитов прочно связывается с кислородом, образуя оксигемоглобин. Однако в периферических тканях эритроциты должны отдать этот кислород другим клеткам. Это осуществляется благодаря явлению, известному под названием эффекта Бора. Эффект Бора создается благодаря двум факторам: протонам и 2,3-бисфосфоглицерату (рис. 18.2).
Протоны вытесняют кислород из оксигемоглобина
Работающим мышцам необходим АТФ, и поэтому в них интенсивно функционирует цикл Кребса. В цикле Кребса образуется диоксид углерода, который проникает в эритроциты и реагирует с водой, образуя угольную кислоту (эту реакцию катализирует карбоангидраза). Угольная кислота самопроизвольно диссоциирует, образуя бикарбонат и протон, что приводит к местному возрастанию концентрации протонов (Н+), т.е. понижению pH. Протоны высвобождают кислород из гемоглобина, и кислород диффундирует из эритроцитов в периферические ткани. Здесь он связывается с миоглобином и переносит кислород в дыхательную цепь, где он используется для синтеза АТФ путем окислительного фосфорилирования.
Высвобождение кислорода в периферических тканях: 2,3-бисфосфоглицерат (2,3-БФГ) стабилизирует структуру дезоксигемоглобина. Помимо протонов, в создании эффекта Бора принимает участие 2,3-бисфосфоглицерат (2.3-БФГ) (рис. 18.2). В медицинской литературе распространено и другое название этого вещества — 2,3-дифосфоглицерат (2,3-ДФГ). 2.3-БФГ образуется в анаэробных условиях в эритроцитах. Процесс образования 2,3-БФГ происходит в дополнительной реакции гликолиза, получившей название
2,3-БФГ-шунта (шунт Рапопорта-Люберинга) (рис. 18.2). В периферических тканях молекула 2,3-БФГ связывается с дезоксигемоглобином и стабилизирует его структуру, не позволяя ему захватывать кислород от соседней молекулы оксигемоглобина.
Связывание гемоглобина с кислородом в легких. Эритроциты переносят дезоксигемоглобин и связанный с ним СО2 в легкие. Поскольку в легких имеет место высокое парциальное давление кислорода, он вытесняет С02 из гемоглобина, и С02 выходит из легких при выдохе. Кислород связывается с гемоглобином, образуя оксигемоглобин, 2,3-БФГ вытесняется, и эритроциты переносят к периферическим тканям следующую порцию кислорода.
Общая характеристика систем энергообеспечения мышечной деятельности.
Энергия, как известно, представляет собой общую количественную меру, связывающую воедино все явления природы, разные формы движения материи
Из всех видов энергии, образующейся и использующейся в различных физических процессах(тепловая, механическая, химическая и др.)применительно к мышечной деятельности, основное внимание должно быть сконцентрировано на химической энергии организма, источником которой являются пищевые продукты и её преобразовании в механическую энергию двигательной деятельности человека
Энергия, высвобождаемая во время расщепления пищевых продуктов, используется для производства аденозинтрифосфата (АТФ), который депонируется в мышечных клетках и является своеобразным топливом для производства механической энергии мышечного сокращения.
Энергию для мышечного сокращения даёт расщепление аденозинтрифосфата (АТФ) до аденозиндифосфата (АДФ) и неорганического фосфата (Ф). Количество АТФ в мышцах невелико и его достаточно для обеспечения высокоинтенсивной работы лишь в течении 1 – 2 с. Для продолжения работы необходим ресинтез АТФ, который производится за счет энерго отдающих реакций трёх типов. Восполнение запасов АТФ в мышцах позволяет поддерживать постоянный уровень его концентрации, необходимый для полноценного мышечного сокращения.
Ресинтез АТФ обеспечивается как в анаэробных, так и в аэробных реакциях с привлечением в качестве энергетических источников запасов креатинфосфата (КФ) и АДФ, содержащихся в мышечных тканях, а также богатых энергией субстратов (гликоген мышц и печени, запасы липозной ткани и др.). Химические реакции, приводящие к обеспечению мышц энергией протекают в трёх энергетических системах:
- анаэробной алактатной
- анаэробной лактатной (гликолитической)
- аэробной.
Образование энергии в первых двух системах осуществляется в процессе химических реакций, не требующих наличия кислорода. Третья система предусматривает энергообеспечение мышечной деятельности в результате реакций окисления, протекающих с участием кислорода. Наиболее общие представления о последовательности включения и количественных соотношениях в энергообеспечении мышечной деятельности каждой из указанных систем приведены на рис. 1.

Рис. 1. Последовательность и количественные соотношения процессов энергообеспечения мышечной деятельности у квалифицированных спортсменов в различных энергетических системах (схема): 1 – алактатной; 2 – лактатной; 3 – аэробной.
Возможности каждой из указанных энергетических систем определяются мощностью, т. е. скоростью освобождения энергии в метаболических процессах, и ёмкостью, которая определяется величиной и эффективностью использования субстратных фондов.
Бескислородный этап энергетического обмена
Бескислородный этап протекает в цитоплазме клеток и в случае аэробных организмов включает только гликолиз — ферментативное многоступенчатое окисление глюкозы и ее расщепление до пировиноградной кислоты, которую также называют пируватом.
Молекула глюкозы включает шесть атомов углерода.
При гликолизе она расщепляется до двух молекул пирувата, который включает три атома углерода. При этом отщепляется часть атомов водорода, которые передаются на кофермент НАД, который, в свою очередь, потом будет участвовать в кислородном этапе.
Часть выделяющейся при гликолизе энергии запасается в молекулах АТФ. На одну молекулу глюкозы синтезируется всего две молекулы АТФ.
Энергия, оставшаяся в пирувате, запасенная в НАД, у аэробов далее будет извлечена на следующем этапе энергетического обмена.
В анаэробных условиях, когда кислородный этап клеточного дыхания отсутствует, пируват «обезвреживается» в молочную кислоту или подвергается брожению.
При этом энергия не запасается. Таким образом, здесь полезный энергетический выход обеспечивается только малоэффектвным гликолизом.
Окисление
Окислению могут подвергаться:
- карбонильная группа
- оба конца углеродной цепи (карбонильная группа и гидроксогруппа у шестого атома углерода)
- гидроксогруппа у шестого атома углерода независимо от карбонильной
Вид окисления зависит от природы окислителя.
Мягкое окисление. Гликоновые кислоты
- Окислитель: бромная вода
- Что окисляется: карбонильная группа альдоз. Кетозы в этих условиях не окисляются и могут быть таким образом выделены из смесей с альдозами.
- Продукты окисления: гликоновые кислоты (из ациклических моносахаридов), пяти- и шестичленные лактоны (из циклических).
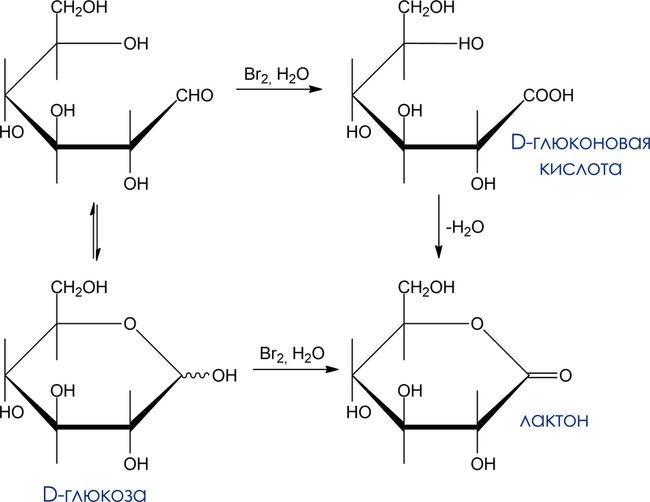
Восстанавливающие моносахариды. Качественная реакция на альдегидную группу
- Окислитель: катионы металлов Ag+ ([Ag(NH3)2]OH – реактив Толенса) и Cu2+ (комплекс Cu2+ с тартрат-ионом – реактив Фелинга) в щелочной среде
- Что окисляется: карбонильная группа альдоз и кетоз
- Продукты окисления: гликоновые кислоты и продукты деструктивного распада
Альдоза + [Ag(NH3)2]+ → гликоновая кислота + Ag + продукты деструктивного окисления
Альдоза + Cu2+ → гликоновая кислота + Cu2O + продукты деструктивного окисления
Восстанавливающие углеводы – углеводы, способные восстанавливать реактивы Толенса и Фелинга. Кетозы проявляют восстанавливающие свойства за счет изомеризации в щелочной среде в альдозы, которые и взаимодействуют далее с окислителем. Процесс превращения кетозы в альдозу происходит в результате енолизации. Образующийся из кетозы енол является общим для нее и 2-х альдоз (эпимеров по С-2). Так, в слабощелочном растворе в равновесии с D-фруктозой находятся ендиол, D-глюкоза и D-манноза.
Эпимеризация – взаимопревращения между альдозами, эпимерами по С2 в щелочном растворе.
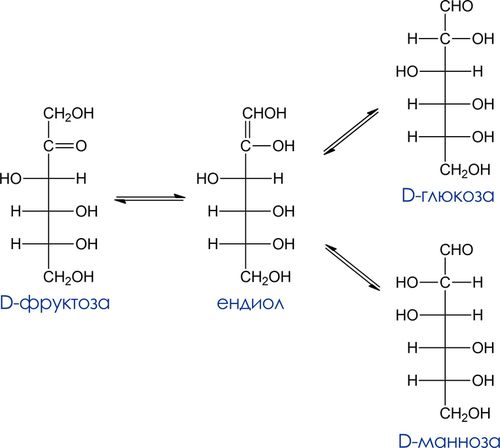
Жесткое окисление. Гликаровые кислоты
- Окислитель: разбавленная азотная кислота
- Что окисляется: оба конца углеродной цепи. Окисление кетоз азотной кислотой протекает с расщеплением С-С связей.
- Продукты окисления: гликаровые кислоты
При образовании гликаровых кислот, происходит “уравнивание” функциональных групп на концах цепи и из некоторых альдоз образуются мезо-соединения.
Ферментативное окисление в организме. Гликуроновые кислоты
- Окислитель: ферменты в организме. В лабораторных условиях для защиты карбонильной группы проводят многостадийный синтез.
- Что окисляется: гидроксогруппа у шестого атома углерода независимо от карбонильной
- Продукты окисления: гликуроновые кислоты
Гликуроновые кислоты входят в состав полисахаридов (пектиновые вещества, гепарин). Важная биологическая роль D-глюкуроновой кислоты состоит в том, что многие токсичные вещества выводятся из организма с мочой в виде растворимых глюкуронидов.
Что такое гликолиз
В общих чертах, гликолиз — это процесс, который происходит в цитоплазме всех клеток: от низших бактерий до высших млекопитающих — людей. Для некоторых это единственный метаболический путь доставки энергии клетке — например, для бактерий, для других же это метаболический путь, в который «сливаются» и другие метаболиты — белки и липиды.
Гликолиз — это основной путь расщепления глюкозы в нашем организме, основное биологическое значениеэтого процесса заключается в получении энергии и других продуктов обмена.
 Гликолиз происходит как в аэробных, так и в анаэробных условиях. Процесс в анаэробных условиях (недостаток кислорода) называется анаэробным гликолизом — конечным продуктом лактата и ферментации в дрожжах — алкогольной или молочнокислой ферментации.
Гликолиз происходит как в аэробных, так и в анаэробных условиях. Процесс в анаэробных условиях (недостаток кислорода) называется анаэробным гликолизом — конечным продуктом лактата и ферментации в дрожжах — алкогольной или молочнокислой ферментации.
При наличии кислорода этот процесс называется аэробным гликолизом — конечным продуктом выступают CO2 и H2O,
Гликолиз представляет собой серию реакций, в которых одна молекула глюкозы распадается на две молекулы пирувата. Но так происходит не всегда, в случаях когда доступ кислорода в этой реакции затруднен, проще говоря, в анаэробных условиях, то пируват превращается в лактат или спирт.
Механизмы анаэробного метаболизма: гликолиз в анаэробных условиях
В анаэробных условиях O2 отсутствует. Отсутствие конечного субстрата O2 ингибирует активность дыхательных структур (до этого они неактивны), и таким образом конечным продуктом распада глюкозы в анаэробных условиях является лактат,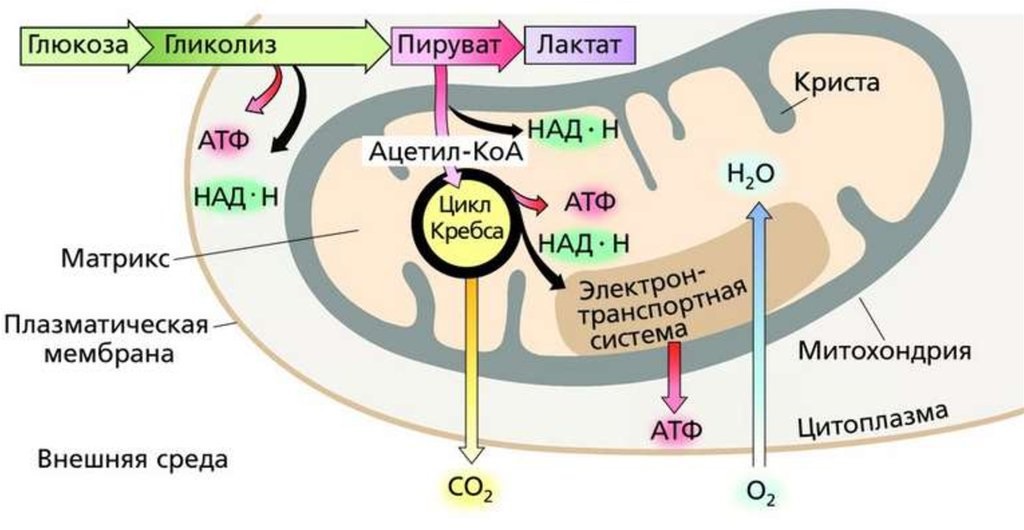
Анаэробный распад глюкозы в физиологических условиях наблюдается в эритроцитах (без митохондрий) и в сильно сокращающихся мышцах (недостаточное кровоснабжение кислородом), это наблюдается при сильной физической работе — присутствует истощение O2 в мышечных клетках (в форме оксимиоглобина).
Лактат в мышечных клетках приводит к мышечной лихорадке, которая преодолевается неинтенсивной мышечной активностью, направленной на полное расщепление лактата в мышцах, до CO2 , H2O и H2 в результате аэробного гликолиза.
При патологических состояниях и онкологических заболеваниях, у многих людей наблюдается так называемый эффект Пастера, при котором клетки из-за уменьшенного поступления крови или кислорода переключаются на анаэробный распад глюкозы, связанный с повышенным ее потреблением (повышенная экспрессия трансмиттеров глюкозы GLUT1 ) и гипогликемией.
Анаэробный гликолиз, как при физиологических, так и при патологических состояниях, характеризуется повышенным уровнем конечного продукта в результате расщепления его лактата в плазме крови — лактатемией, что может привести к ацидозу.
Дальнейшее расщепление лактата
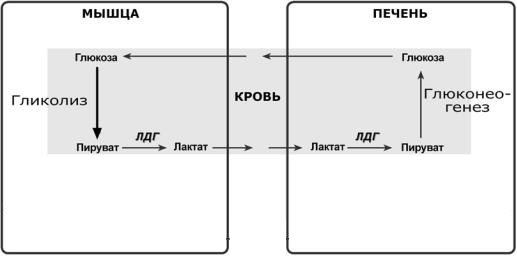
Лактат метаболизируется из печени в пируват, катализируя реакцию в обратном направлении, и глюкоза синтезируется пируватом с использованием ферментов глюконеогенеза в печени через цикл Кори (лактатный цикл).
Вот в этой схеме и кроется ответ на вопрос о том, как влияет анаэробный гликолиз на производство энергии, ведь мы видим, что наша молочная кислота, образовавшаяся в мышцах, через кровь попадает в печень и участвует в процессе глюконеогенеза — создания глюкозы из неуглеводных продуктов.
Энергетический обмен
Некоторые облигатные анаэробы используют брожение , другие — анаэробное дыхание . Аэротолерантные организмы строго ферментируют. В присутствии кислорода факультативные анаэробы используют аэробное дыхание ; без кислорода некоторые из них ферментируют; некоторые используют анаэробное дыхание.
Ферментация
Существует множество анаэробных ферментативных реакций.
Ферментативные анаэробные организмы в основном используют путь молочнокислого брожения:
- C 6 H 12 O 6 + 2 АДФ + 2 фосфата → 2 молочная кислота + 2 АТФ + 2 H 2 O
Энергия, выделяемая в этой реакции (без АДФ и фосфата), составляет примерно 150 кДж на моль , что сохраняется при образовании двух АТФ из АДФ на глюкозу . Это всего лишь 5% энергии на молекулу сахара, которую генерирует типичная аэробная реакция с использованием высокой энергии O 2 .
Растения и грибы (например, дрожжи) обычно используют спиртовое (этанольное) брожение, когда кислород становится ограниченным:
- C 6 H 12 O 6 ( глюкоза ) + 2 ADP + 2 фосфата → 2 C 2 H 5 OH + 2 CO 2 ↑ + 2 ATP + 2 H 2 O
Выделяемая энергия составляет около 180 кДж на моль, что сохраняется при выработке двух АТФ из АДФ на глюкозу.
Анаэробные бактерии и археи использовать эти и многие другие ферментативные пути, например, кислоты , пропионовой брожения, масляная кислота брожения, ферментации растворителя, кислоты брожения смешивают, бутандиол ферментации , Stickland ферментации , acetogenesis или Метаногенез .
Стадия реакции окисления глюкозы
- Первый этап. На первом этапе происходит фосфорилирование глюкозы. Активирование сахарида происходит путем фосфолирирования по шестому углеродному атому.
- Второй этап. Происходит процесс изомеризации глюкозы-6-фосфата. На данном этапе глюкоза обращается во фруктозу-6-фосфат под действием каталитического фосфоглюкоизомераза.
- Третий этап. Фосфорилирование фруктозы-6-фосфата. На данном этапе происходит образование фруктозо-1,6-дифосфата (называемого также альдолазой) под воздействием фосфофруктокиназы-1. Она участвует в сопровождении фосфорильной группы от аденозинтрифосфорной кислоты до молекулы фруктозы.
- Четвертый этап. На данном этапе происходит расщепление альдолазы. В результате образуются две молекулы триозофосфата, в частности кетозы и эльдозы.
- Пятый этап. Изомеризация триозофосфатов. На данном этапе происходит отправка глицеральдегид-3-фосфата на следующие этапы глюкозного расщепления. При этом происходит переход дигидроксиацетонфосфата в форму глицеральдегид-3-фосфата. Данный переход осуществляется под действием ферментов.
- Шестой этап. Процесс окисления глицеральдегид-3-фосфата. На данном этапе происходит окисление молекулы и ее последующее фосфорилирование до дифосфоглицерата-1,3.
- Седьмой этап. Данный этап предполагает перенос из 1,3-дифосфоглицерата фосфатной группы на АДФ. В конечном результате этого этапа образуется 3-фосфоглицерат и АТФ.
Теги
окисление глюкозы.окисления глюкозы окисление глюкозы. Глюкоза можетокисления глюкозы.окисления глюкозы.окисление глюкозы происходитокислении глюкозы образуетсяокисление глюкозы.шестиуглеродной глюкозы на Гликолиз Гликолиз представляетПроцесс гликолиза происходитпроцесс гликолиза можетвид гликолиза называется Гликолиз анаэробноготаком гликолизе окислениеанаэробный гликолиз менееанаэробного гликолиза вМеханизм гликолиза глюкозы. Реакция окисленияхимическая реакция окисленияхимическая реакция возможнагликолитической реакции необходимопромежуточной реакции субстратногоэтой реакции являетсяв АТФ ив АТФ запасаетсяи АТФ.в АТФ.
пируваталактатаброжениемрольрегуляцияхимияэритроцитахкрахмалэнергетическаяраствороврисконцентрациитолькоспиртовскорости
Значение гликолиза
Гликолиз — это уникальный, однако не единственный вид окисления глюкозы в организме, клетке как прокариотов, так и эукариотов. Ферменты гликолиза являются водорастворимыми. Реакция гликолиза в некоторых тканях и клетках может происходить только таким образом, например, в мозгу и клетках нефронов печени. Другие способы окисления глюкозы в этих органах не используются. Однако не везде функции гликолиза одинаковы. Например, жировые ткани и печени в процессе пищеварения добывают необходимые субстраты из глюкозы для синтеза жиров. Многие растения используют гликолиз как способ добычи основной части энергии.






